






ご挨拶
海洋においては、外洋域、深海、海底熱水孔、極地、赤潮海域、沿岸養殖漁場など種々の特徴を有する環境が存在し、そこには極めて多様な微生物が生息している。これら海洋並びに湖沼に生息する微生物(ファージ、細菌、古細菌および真核微細藻)の多様性と水圏環境下での生存戦略や他の生物との相互作用、それに関る物質の作用機作を生態学、生理・生化学、分子生物学を総合して解析するとともに、それらがもつ新規な遺伝子資源を利用することを目指している。これを「海洋分子微生物学」と位置づけている。
新着情報
- 2024.4.5
-
新年度の顔合わせを行いました。今年度は新たに4回生4名を迎えました。これからよろしくお願いします。

- 2024.3.27
-
本研究室から8名が卒業を迎えました。新天地での活躍をお祈り致しております。

- 2024.3.14
-
冷え込みが厳しい冬場を抜け、春の気配が感じられるようになりました。吉田山でランチピクニックを行いました。

- 2024.3.12
-
システム移行のためしばらくHPが閲覧できない状態となっておりました。ご心配・ご迷惑をおかけしました。
- 2024.3.8
-
3月6~8日に沖縄科学技術大学院大学のFilip Husnik 准教授の研究室を訪問しました。本研究室の神川准教授が非光合成藻類の進化について講演を行い、意見交換しました。空き時間には研究室や大学の設備を見学し、教員や学生と議論を深めました。


- 2024.3.5
-
本研究室OBの武部博士(R4学位取得)と富永博士(R2学位取得)を筆頭著者および共著者とする論文がmSystems誌に受理されました。
Taxonomic difference in marine bloom-forming phytoplanktonic species affects the dynamics of both bloom-responding prokaryotes and prokaryotic viruses
Takebe, H., Tominaga, K., Isozaki, T., Watanabe, T., Yamamoto, K., Kamikawa, R. and Yoshida, T.
mSystems. 2024. doi: 10.1128/msystems.00949-23.
- 2024.2.13
-
2024年2月13日、課題研究発表会が行われました。当研究室から2名がポスター発表を行いました。
ポスター発表 題目
江端康太 海洋におけるナノ真核藻類が原核微生物群集に与える影響に関する研究 筈井優斗 ハプト藻類Pavlomulina ranunculiformis における内在性ウイルスの探索 2月13日、歓送迎会および謝恩会を行いました。
来年度から研究室に加わるメンバーも交え、歓談を楽しみ、研究室を卒業される方々の門出をお祝いしました。皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2024.2.6
-
2024年2月6日、修士論文発表会が行われました。当研究室から5名が口頭発表を行いました。
口頭発表 題目
久保さやか 定量PCR法および可視超分光法による微細藻におけるウイルス感染細胞の検出 山田隆太郎 集積メタオミクスによる大阪湾におけるナノピコ真核生物の多様性解析 小出斎 Ecophysiological characteristics of coexisting bacteria of the haptophyte Pavlomulina ranunculiformis (ハプト藻類Pavlomulina ranunculiformisの共存真正細菌の生理生態学的特性) 嶋田理香子 海洋未培養細菌SAR86系統の大阪湾における優占個体群の多様性と動態 鈴木晟太 淡水底泥からの水素生成型一酸化炭素酸化菌の分離及びゲノム性状解析
- 2024.1.18
-
博士後期課程3年の西田志穂さんの博士論文申請講演会が行われました。
題目
西田志穂 Studies on microbial diversity of carbon monoxide utilizers in the rare biosphere of aquatic environments (⽔圏の希少⽣物圏に潜在する⼀酸化炭素利⽤菌の多様性に関する研究)
- 2024.1.9
-
吉田教授と東京大学でポスドクをされている本研究室OBの富永博士(R2学位取得)が共著者として参画した論文がmSystems誌に受理されました。
Virus–prokaryote infection pairs associated with prokaryotic production in a freshwater lake
Shen, S., Tominaga, K., Tsuchiya, K., Matsuda, T., Yoshida, T. and Shimizu, Y.
mSystems. 2024. doi: 10.1128/msystems.00906-23
- 2023.12.26
-
東京大学でポスドクをされているOB富永博士(R2学位取得)を筆頭著者とする論文がEnvironmental Microbiology Reports誌に受理されました。
Population-level prokaryotic community structures associated with ferromanganese nodules in the Clarion-Clipperton Zone (Pacific Ocean) revealed by 16S rRNA gene amplicon sequencing
Tominaga, K., Takebe, H., Murakami, C., Tsune, A., Okamura, O., Ikegami, I., Onishi, Y., Kamikawa, R. and Yoshida, T. Environ. Microbiol. Rep. 2023; 1-13. doi: 10.1111/1758-2229.13224
- 2023.12.25
-
本年度も当研究室OB・OGの皆様からお歳暮を頂きました。研究室一同、ありがたく賞味させていただきました。OB・OGの皆様、お心遣いいただきありがとうございました。

- 2023.12.22
-
当研究室の前教授である左子芳彦先生が来研され、歓談の後、記念写真を撮影しました。久しぶりの再会に研究室も盛り上がりを見せ、非常に楽しい時間となりました。今後ともよろしくお願いいたします。

- 2023.12.19
-
宇都宮大学の佐藤博士(元京都大学農学研究科博士課程学生)を筆頭著者とする論文がApplied and Environmental Microbiology誌に受理されました。吉田教授と本研究室OBである武部博士と富永博士が最終著者および共著者として参画しています。
A rumen virosphere with implications of contribution to fermentation and methane production, and endemism in cattle breeds and individuals
Sato, Y., Takebe, H., Tominaga, K., Yasuda, J., Kumagai, H., Hirooka, H. and Yoshida, T. Appl. Environ. Microbiol. 2024. doi: 10.1128/aem.01581-23
- 2023.12.9
-
2023年12月9日、近畿大学農学部において令和5年度水産学会近畿支部後期例会が開催されました。M2の久保さやかさん、M1の百合岡咲紀さんが口頭発表を行い、M2の小出斎さん、鈴木晟太さん、M1の光浪健太さんがポスター発表を行いました。
口頭発表 題目
久保さやか ハイパースペクトルカメラを用いたウイルス感染藻類細胞検出の試み 百合岡咲紀 大阪湾において周期的に出現する優占緑藻類の発見 ポスター発表 題目
小出斎 ラピ藻Pavlomulina ranunculiformis NIES-3900株の共存真正細菌の生理生態学的特性 鈴木晟太 淡水底質環境から分離された水素生成型一酸化炭素酸化菌のゲノム性状解析 光浪健太 大阪湾に優占する古細菌系統Marine Group II Euryarchaeotaの季節動態解析
- 2023.11.30
-
2023年11月28~30日、アクトシティ浜松コングレスセンターにおいて日本微生物生態学会第36回大会が開催されました。D3の西田志穂さん、D2の今浦由就さん、M2の久保さやかさん、小出斎さん、鈴木晟太さん、M1の佐野圭志郎さん、武村拓海さん、光浪健太さん、宮浦緋里さん、百合岡咲紀さんがポスター発表を行いました。

題目
西田志穂 琵琶湖湖底に潜在する水素生成型一酸化炭素利用菌の探索と分離 今浦由就 「一酸化炭素を食べる」海洋原核生物群集の大阪湾における季節的動態 久保さやか ハイパースペクトルカメラを用いた微細藻類におけるウイルス感染細胞検出の試み 小出斎 ハプト藻類Pavlomulina ranunculiformis NIES-3900株の共存真正細菌の生理生態学的特性 鈴木晟太 淡水底質環境から分離された水素生成一酸化炭素酸化菌のゲノム・生理性状解析 佐野圭志郎 大阪湾に優占するピコ真核藻類の生理生態学的特性解明に向けたメタオミクス解析 武村拓海 海洋ブルーム形成真核微細藻類Heterosigma akashiwoに感染するウイルスHaVの感染過程の解明 光浪健太 大阪湾に優占する古細菌系統Marine Group II Euryarchaeota の季節動態の解明 宮浦緋里 ハプト植物門ラピ藻綱に属する真核微細藻類の分布と環境要因との関係 百合岡咲紀 大阪湾において周期的に出現する優占緑藻類の発見
- 2023.11.20
-
11月20日、真如堂で記念写真を撮影しました。紅葉の色彩が美しく染まった風景の中で、心地よいひとときを過ごしました。

- 2023.11.19
-
2023年11月18~19日、京都大学農学部において日本共生生物学会第7回大会が開催されました。
特定研究員の石井悠さんが口頭発表を行いました。
題目
石井悠 サンゴ共生藻の光合成産物の資源分配を介した生活スタイルの多様化
- 2023.10.2
-
10月2日、特定研究員の石井悠さんが日本動物学会女性研究者奨励OM賞を受賞されたことをお祝いし、皆でケーキを頂きました。
石井さんのこの度のご受賞を、心よりお祝い申し上げます。

- 2023.9.7
-
博士課程の西田さんの論文が Archives of Microbiology誌に受理されました。
Construction of multiple metagenome assembled genomes containing carbon monoxide dehydrogenases from anaerobic carbon monoxide enrichment cultures
S Nishida, K Omae, M Inoue, Y Sako, R Kamikawa, T Yoshida, Archives of Microbiology 205 (8), 292, 2023.一酸化炭素 (CO) は微生物を含む多くの生物にとって有毒ですが、微生物の中にはCOをエネルギー源および炭素源として利用可能なCO利用菌が存在することが報告されてきました。中でも水素生成型CO利用菌は環境中からCOを除去し他の微生物にとってエネルギー源となる水素を供給することから、生態学的に重要であると考えられてきました。しかし、水素生成型CO利用菌は環境中での相対存在量が極めて低く、希少生物圏 (rare biosphere) の構成種であると推測され、環境における本菌の研究は困難です。そこで本研究ではあらかじめ環境試料にCOを封入して水素生成型CO利用菌を集積しておいた培養試料由来のDNAをメタゲノム解析に供することで、水素生成型CO利用菌のゲノムを獲得し、そのCO利用能を明らかにすることを目的としました。メタゲノム解析の結果、高品質な水素生成型CO利用菌のゲノムを2つ得ることに成功し、それらが既知のCO利用能の他に新規なCO利用能を有する可能性を見出しました。本研究によって、集積培養とメタオミクス解析の統合手法がrare biosphereの構成種である水素生成型CO利用菌の研究において効果的な研究手法であることを証明しました。
- 2023.08.06
-
先日、本研究室の神川准教授が誕生日を迎えられました。それに際し、研究室一同より日ごろの感謝を込めて、日本酒をプレゼント致しました。神川先生、おめでとうございます。

- 2023.07.31
-
2023年7月31日、博士後期課程の西田志穂さん、今浦由就さん、修士課程の鈴木晟太さんが執筆した論文が受理されたお祝いに、皆でケーキを頂きました。お三方とも、この度は誠におめでとうございます。


- 2023.7.24
-
7月24日、京都下鴨神社の御手洗祭に参加しました。
無病息災を祈願し、身も心もリフレッシュしました。


- 2023.6.27
-
修士課程の鈴木晟太さんが執筆した論文が Microbiology Resource Announcements 誌に受理されました。
Draft Genome Sequence of Thermolongibacillus altinsuensis Strain B1-1, a Novel Hydrogenogenic CO Oxidizer Isolated from Sediment from Lake Biwa in Japan
Jota Suzuki, Yoshinari Imaura, Shiho Nishida, Ryoma Kamikawa, Takashi Yoshida(2023), Microbiol Resour Announc. Jun 5:e0033423. doi: 10.1128/mra.00334-23.淡水湖である琵琶湖の堆積物から、通性嫌気性好熱性水素発生CO酸化細菌B1-1株が分離されました。本株は水素生成CO酸化能を有するThermolongibacillus altinsuensisの新規株であり、ドラフトゲノム配列内には水素生成型CO酸化菌が有するcoo-ech遺伝子クラスターが認められました。
- 2023.5.9
-
博士後期課程の今浦由就さんが執筆した論文が Applied and Environmental Microbiology 誌に受理されました。
Isolation, genomic sequence and physiological characterization of Parageobacillus sp. G301, an isolate capable of both hydrogenogenic and aerobic carbon monoxide oxidation
Yoshinari Imaura, Shunsuke Okamoto, Taiki Hino, Yusuke Ogami, Yuka Adachi Katayama, Ayumi Tanimura, Masao Inoue, Ryoma Kamikawa, Takashi Yoshida, Yoshihiko Sako (2023), Appl. Environ. Microbiol. (In press).一酸化炭素 (CO) 酸化菌は、多くの生物に有毒なCOをエネルギー源として利用します。CO酸化菌が有するCO酸化の鍵酵素であるCOデヒドロゲナーゼ (CODH)は、タンパク質ファミリーの異なる2タイプに大別され、両者は共役する呼吸機構や機能可能な酸素濃度が異なります。これまでの研究に用いられてきたCO酸化菌はいずれか一方のCODHのみをもつため、それぞれのCO酸化菌がCOを利用可能な条件は限られていると考えられていました。本研究では、両タイプのCODHをもつ真正細菌分離株 Parageobacillus sp. G301 株を報告し、これまでにない幅広い条件でCO酸化が可能であることを示しました。大規模な比較ゲノム解析により、本菌が同様の生理学的特徴を有する世界で唯一の分離株であることを明らかにするとともに、CODH遺伝子の保有がゲノム性状や細胞機能に与える影響について考察しています。
- 2023.4.24
-
システム移行のためHPの更新が途絶えておりました。ご心配・ご迷惑をおかけしました。
- 2023.4.21
-
本研究室のOBでJAMSTECの研究員である福山さんにご講演頂きました。
短い時間でしたが興味深いお話をお聞かせいただき、大変勉強になりました。
お土産も頂き、美味しくいただきました。ありがとうございました。

- 2023.4.3
-
新年度の顔合わせを行いました。
今年度は新たに4回生2名、修士3名を迎えました。これからよろしくお願いします。
- 2023.3.24
-
3月24日、本研究室から6名が卒業を迎えました。新天地での活躍をお祈り致しております。

- 2023.3.22
-
エクス=マルセイユ大学Pascal Hingamp先生が 8年ぶりに来日されました。メタゲノム解析をご専門とされています。
研究の近況に加え、琵琶湖を視察しながら、コロナ禍の国内状況や教育状況について意見交換をし、リラックスした楽しいひと時となりました。
- 2023.2.14
-
2月10日、歓送迎会および謝恩会を行いました。
来年度から研究室に加わる方も交えて歓談を楽しみ、研究室を卒業される方々の門出をお祝いしました。
皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2023.2.14
-
2023年2月10日、修士論文発表会が行われました。当研究室から5名が口頭発表を行いました。
口頭発表 題目芦澤柊人 メタオミクス解析を用いた大阪湾に優占する未培養古細菌Marine Group II Euryarchaeotaの季節動態とウイルスとの相互作用の解明 芦谷朋香 Diel infection dynamics of multiple viruses infecting the dominant cyanobacterium in Osaka Bay revealed by the multiple-omics analyses
(マルチオミクス解析を用いた大阪湾における優占シアノバクテリア感染性ウイルスの日周感染動態の解明)今髙里莉 全層循環過程を通じた琵琶湖第二湖盆の堆積物表層における細菌叢解析と一酸化炭素酸化菌の探索 中村明美 大阪湾における優占ピコ緑藻とウイルスとの相互作用 山本葵 新規Nitzschia属珪藻のオルガネラゲノミクス及びトランスクリプトミクスによる非光合成性葉緑体進化の解明 2023年2月14日、課題研究発表会が行われました。当研究室から5名がポスター発表を行いました。
ポスター発表 題目佐野奎志郎 メタオミクス解析による大阪湾に優占するピコ真核藻類のオルガネラゲノムの決定と日周動態の解明 武村拓海 定量的 PCR 法による海洋性有害赤潮藻類 Heterosigma akashiwo に感染するウイルス HaV の増殖過程の解明 浜名俊輔 海洋ヴィローム解析による高精度宿主予測とウイルス-宿主間相互作用解析 宮浦緋里 月光が真核微細藻類Emiliania huxleyi の分裂および色素組成変化の周期性に与える影響の解明 迎井慎太朗 比較トランスクリプトーム解析によるマングローブ林河口由来非光合成性珪藻における塩濃度変動に対する適応の解明
- 2023.1.5
-
東京大学でポスドクをしているOB富永さん(R2学位取得)の論文がmSystems誌に受理されました。
Prevalence of viral frequency-dependent infection in coastal marine prokaryotes revealed using monthly time series virome analysis.
Tominaga,K., Ogawa-Haruki, N., Nishimura, Y., Watai, H., Yamamoto, K., Ogata, H. and Yoshida, T.,
mSystems 2023, DOI: https://doi.org/10.1128/msystems.00931-22ウイルス感染は地球上のあらゆる生物に起きると考えられます。しかし、海洋の優占原核生物は分離することさえ難しく、本当にウイルスに感染が起きているのか検証するのは困難でした。本研究では、大阪湾の原核生物とウイルスの動態をメタゲノム解析により約2年間月ごとに追跡し、ゲノム情報に基づくウイルスの宿主予測法と組み合わせることで、優占原核生物とそのウイルスの間で多数の共起関係を見いだしました。この結果は、原核生物の分類群や季節動態に関係なく、優占種であればいずれもウイルス感染が起こることを示唆しています。本研究により、原核生物優占種のウイルス感染による溶菌は、海洋物質循環や群集多様性の維持に大きく寄与するプロセスであることが支持されました。
詳細はこちらから https://www.kyoto-u.ac.jp/ja/research-news/2023-02-24-4
- 2023.1.5
-
高知大学でポスドクをされているOB森本さん(H30博士学位取得)の論文が Applied and Environmental Microbiology誌に受理されました。
Ecological dynamics of broad- and narrow-host-range viruses infecting the bloom-forming toxic cyanobacterium Microcystis aeruginosa.
Morimoto,D., Yoshida, N., Sasaki, A., Nakagawa, S., Sako,Y. and Yoshida, T.
Applied and Environmental Microbiology, 89, e02111-22 , 2023. DOI: https://doi.org/10.1128/aem.02111-22アオコ原因ラン藻ミクロキスティスは、多種多様なウイルスと感染を巡る攻防を繰り広げています。本研究では、環境において宿主域の異なるウイルスが、本種に与える影響を調べました。その結果、広宿主域ウイルスの感染により本種の種内個体群の比率は変動するが、全体の生物量は増加・維持されることが示されました。この両者の共存の背景には、本種の卓越したウイルス耐性機構が介在していることが明らかとなりました。
詳細はこちらからhttps://www.kyoto-u.ac.jp/ja/research-news/2023-02-24-5
- 2022.12.22
-
当研究室2017年度修了 (修士) の大井さんと2017年度卒業 (学士) の鈴木さんが来研されました。 吉田教授、在校生を交えて昔懐かしい話に花を咲かせました。

- 2022.12.21
-
当研究室の前教授である左子芳彦名誉教授が来研され、歓談の後記念撮影をしました。久しぶりの再会に研究室も盛り上がりを見せ、非常に楽しい時間となりました。今後ともよろしくお願いいたします。

- 2022.12.15
-
博士後期課程3年の武部紘明さんの博士論文申請講演会が開かれました。 OBも多数来研され、大盛況の中行われました。
題目
武部紘明 Molecular ecological studies on effects of a marine harmful algal bloom-forming species, Heterosigma akashiwo, on prokaryotic community (海洋性有害赤潮形成藻類Heterosigma akashiwoが原核生物群集に 及ぼす影響に関する分子生態学的研究) 
- 2022.12.3
-
2022年12月3日、令和四年度水産学会近畿支部後期例会が開催されました。 M2の芦谷朋香さんが口頭発表を行いました。
題目
芦谷朋香 大阪湾における優占シアノバクテリア感染性ウイルスの日周感染動態の解明
- 2022.11.14
-
11月14日、真如堂で記念写真を撮影しました。
美しく色づいた紅葉の帳の中、和やかなひと時を過ごしました。

- 2022.11.9
-
2022年11月9日、神戸ポートピアホテルにおいてThe 9th International Human Microbiome Consortium Congress 2022が開催されました。 D1の片山(足立)夕花さんがポスター発表を行いました。
題目
片山(足立)夕花 Comprehensive genomic analysis uncovered CO-utilizing pathways in human gut microbiome
- 2022.10.31~11.3
-
2022年10月31日~11月3日、札幌コンベンションセンターにおいて日本微生物生態学会第35回大会が開催されました。 D3の武部紘明さん、D1の今浦由就さんが口頭発表を、D2の西田志穂さん、M2の芦谷朋香さん、今高里莉さん、M1の小出斎さん、鈴木晟太さんがポスター発表を行いました。

題目
武部紘明 海洋性真核微細藻類に対するウイルス感染が原核生物群集動態に及ぼす影響 今浦由就 琵琶湖第二湖盆底泥における潜在的CO酸化活性能を持つ微生物の探索 西田志穂 水圏環境中に潜在する水素生成型一酸化炭素酸化菌の探索とゲノム特性の解明 芦谷朋香 大阪湾における優占シアノバクテリア感染性ウイルスの日周感染動態の解明 今高里莉 琵琶湖第二湖盆底泥における潜在的CO酸化活性能を持つ微生物の探索 小出斎 ハプト藻類 Pavlomulina ranunculiformis NIES-3900 株における共存原核生物群集との相互作用 鈴木晟太 淡水底質環境からの水素 (H2) 生成型一酸化炭素 (CO) 酸化菌の分離および性状解析
- 2022.10.24
-
10月24日、定期サンプルなどで日頃大変お世話になっている、大阪府水産技術センターの皆様が採水口をバージョンアップしてくださいました。
研究室一同さらによいサンプルが取れると歓喜しております。ありがとうございました。
- 2022.9.21
-
宮城教育大学で日本学術振興会特別研究員PDをされている石井悠先生をお招きし、講義を行っていただきました。
サンゴと褐虫藻の共生関係の分子メカニズムを明らかにするために先生が行われた実験や遺伝子解析についてお話し頂きました。短い時間でしたが非常に興味深い講義でした。お土産も頂き、大変おいしくいただきました。ありがとうございました。
- 2022.9.14,15
-
9月14~15日、今年度初の大阪湾の24時間サンプリングを行いました。
3時間おきに海水中の微生物群集を真核生物画分、原核生物画分、ウイルス画分に分けて採集するという気を遣う作業でしたが、2部制を導入し協力してスムーズに作業を進めることができました。
疲労困憊での帰路となりましたが、貴重な経験とサンプルを得ることができました。今後、本サンプルを使って面白いデータを取っていこうと思います。

- 2022.9.12
-
9月12日、琵琶湖にてサンプリングが行われました。
38℃と、秋口とは思われない気温の中のサンプリングでしたが、水分補給も挟みながら、何とか乗り切りました。
又、本サンプリングにて、一旦琵琶湖での定期サンプリングは終了となります。琵琶湖調査船「はす」への乗船の許可、及びサンプリングに協力してくださいました、京都大学生態学研究センターの合田様、赤坂様、一年間誠にありがとうございました。
- 2022.8.1
-
OB,OGの皆様から、たくさんのお中元をいただきました。
研究室一同でありがたく賞味させていただきました。
OB・OGの皆様、お心遣い頂きありがとうございました。
- 2022.8.1
-
8月1日、本日は琵琶湖にて、サンプリングが行われました。
行きしなの温度計では、37℃、38℃など、体温をも超える暑さとなり、かなり心配していましたが、琵琶湖ではそよ風が心地よく、何とか乗り切れました。
- 2022.7.24
-
7月24日、本日は京都下賀茂神社で御手洗まつりが行われました。
熱い天気の中、キンキンに冷えた水が心地よく、身も心も現れました!
- 2022.7.20
-
7月20日、R3年卒の星野さんが本研究室にご来訪されました。久しぶりの再会に研究室も盛り上がりを見せ、非常に楽しい時間となりました。
今後ともよろしくお願いいたします。
- 2022.7.12,13
-
7月12~13日、本研究室のOBである高知大学の長崎教授をお招きし、講義を行っていただきました。
長崎先生の研究者人生から始まり、微生物学の推移、最新のウイルス学に至るまで、講義内容は非常に幅広く、更に、直筆イラストでの説明など、濃い、わかりやすい、そして何より面白い講義でした。
本当にありがとうございました。

- 2022.5.26
-
5月26日、海洋深層に存在する微生物、ウイルスを調査するために高知県室戸市の高知県海洋深層水研究所にて京都大学化学研究所の緒方研究室と合同で海洋深層水をサンプリングしました。
当日は夕刻から土砂降りの大雨でしたが、研究所の皆様のご協力のもとで雨を避けて無事サンプルの採取と処理を行えました。

- 2022.5.20
-
5月20日、琵琶湖で湖水および堆積物のサンプリングを行いました。
曇り空でしたが風も波もほとんどなく、心地よい初夏の風の中で、安全、快適に採取を終えることができました。

- 2022.4.20
-
4月20日、大阪湾で新入生にとって初のサンプリングを行いました。
慣れない作業でしたが水産技術センターの大美様、山本様のご協力のもと円滑に進めることができました。
天候にも恵まれ、研究生活のよいスタートを切れました。

- 2022.4.12
-
4月12日、琵琶湖にて水素生成型一酸化炭素酸化菌の探索を目的とした環境試料のサンプリングを行いました。
先月とは打って変わって比叡山に積もる雪も消え、そよ風が心地よい夏日のサンプリングとなりました。
今回より新しく導入された器具により、更に手際よく作業を進めることができました。

- 2022.3.8
-
3月8日、琵琶湖にて水素生成型一酸化炭素酸化菌の探索を目的とした環境試料のサンプリングを行いました。
北西の微風予報にもかかわず、南湖ではしっかり強風、サンプリング地点への道中も、うねりが見られるなど、サンプリング自体が危ぶまれておりました。
しかし、北湖においては、風は微風で安定しており素晴らしいサンプリング日和でした。

- 2022.2.24
-
2022年2月24日、歓送迎会を行いました。
研究室を卒業される方々、来年度から研究室に加わるメンバーも交え、換気に気を配り距離を保ちながら歓談を楽しみ、研究室を卒業される方々へのはなむけとしました。
皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。
- 2022.2.16
-
2月10日、琵琶湖にて水素生成型一酸化炭素酸化菌の探索を目的とした環境試料のサンプリングを行いました。 大学から琵琶湖までの道には雪が残っているなど、寒さが厳しい中でのサンプリングではありましたが、協力し手際よく採取を進めることができました。


- 2022.1.31
-
昨春まで研究員として在籍されていた井上さんの論文が Extremophiles 誌に受理されました。
Biome-specific distribution of Ni-containing carbon monoxide dehydrogenases.
Inoue, M., Omae, K., Nakamoto, I., Kamikawa, R., Yoshida, T. and Sako, Y.,
Extremophiles, 26, 9, 2022.,10.1007%2Fs00792-022-01259-y.一酸化炭素 (CO) 資化菌と呼ばれる微生物は有毒ガスである CO を食べるというユニークな性質を有しています。しかし、この地球のどこに (Where?) どんな (What?) CO 資化菌がいるのかよく分かっていませんでした。 本論文では、環境メタゲノム情報を用いて本菌の鍵酵素遺伝子の生物圏分布を解析しました。その結果、水圏環境やヒト腸内など従来考えられていたよりも幅広い環境において、環境ごとに異なる多種多様な CO 資化菌が分布していることが明らかになりました。
- 2022.1.17
-
当研究室OB・OGの方からお歳暮を頂きました。研究室一同、オミクロン株に注意しつつ大変美味しくいただきました。OB・OGの皆様、お心遣いいただきありがとうございました。


- 2021.12.7
-
2021年12月5日に、令和3年度日本水産学会近畿支部例会が開催されました。修士課程2年の今浦由就さんが口頭発表を行ない、修士課程区分で優秀賞を受賞しました。おめでとうございます!題目
今浦由就 常温湖底堆積物より分離された好熱性Parageobacillus属細菌の一酸化炭素利用能分析 受賞コメント:ご指導、ご尽力いただいた先生方や諸先輩方、研究室の皆様に感謝いたします。今回の受賞を励みとし、残りの修士課程の研究と、博士課程での研究に邁進します。(今浦)
- 2021.11.24
-
廣岡先生のグループと進めてきた黒毛牛のルーメンに関する論文が FEMS Microbiology Ecology誌に受理されました。D3佐藤さんの学位論文の重要なパートでもあります。おめでとうございます。研究内容はもちろんのこと、佐藤さんの執筆、対応力は当ラボの学生にとっても大変刺激になりました。
- 2021.11.24
-
11月16日、先月に引き続いて琵琶湖に行き、水素生成型一酸化炭素酸化菌の探索を目的とした環境試料のサンプリングを行いました。風の影響を心配していましたが、テンポよく作業を進め、安全に採取を終えることができました。培養の結果も出始めており、今後が楽しみなサンプリングとなりました。


- 2021.10.27
-
かなり遅れてご報告いたしますが、今年度新たに2件の研究プロジェクトが採択されました。 科学研究費基盤研究(S)(2021~2025)が採択されました。研究課題は、「ウイルス駆動の海洋低次生態系の時計仕掛けと海洋への影響解明 」です。本プロジェクトでは特定研究員を公募中です。ご興味がある方は下記メールアドレスまで遠慮なくご連絡ください。 2021年度公益財団法人研究所大型研究助成では「始原的呼吸経路を繋ぐ一酸化炭素酸化菌コレクションの構築とその応用基盤」という課題が採択されました。 自画自賛しますが、おめでとうございます。これまでのラボの先輩や共同研究者の方々とのコラボで得た成果を活かし、一丸となって研究を進めていきましょう! HPの研究内容も近くアップしますのでお待ちください。
- 2021.10.26
-
10月19日、水圏堆積物環境における水素生成型一酸化炭素酸化菌の探索を目的に、琵琶湖においてサンプリングを行いました。 琵琶湖の深部は季節的に嫌気的環境となり、有機物から発生する一酸化炭素を用いた嫌気的代謝が起こっている可能性があります。 サンプリングでは京都大学生態学研究センターの合田様、赤塚様にご協力いただき、湖底の環境試料を採取することができました。
- 2021.09.10
-
先日、本研究室の吉田先生がお誕生日を迎えられました。それに当たって、研究室一同よりささやかながら贈り物をお渡ししました。吉田先生、いつもありがとうございます!そしてお誕生日おめでとうございます!

- 2021.08.11
-
先日、本研究室の神川先生が誕生日を迎えられました。それに際し、研究室一同より、日ごろの感謝を込めて、日本酒などをプレゼント致しました。神川先生、おめでとうございます!


- 2021.08.06
-
本年度も当研究室OB・OGの皆様からお中元を頂きました。早速ながら研究室一同、おいしく頂戴いたしました。OB・OGの皆様、いつもお心にかけていただき心より感謝申し上げます。

- 2021.04.06
-
先日、新年度の顔合わせを兼ねて、吉田山でランチピクニックを行いました。今年度は新たに4回生4名、修士2名を迎えました。これからよろしくお願いします。

- 2021.03.27
-
20年以上前からDNAとしてのみ存在が知られていた謎の生物ラピモナズ(Rappemonads)の正体を解明することに成功し、本研究成果はCurrent Biologyのオンライン速報版に掲載されました。
Rappemonads are haptophyte phytoplankton Masanobu Kawachi, Takuro Nakayama, Motoki Kayama, Mami Nomura, Hideaki Miyashita, Othman Bojo, Lesley Rhodes, Stuart Sym, Richard N. Pienaar, Ian Probert, Isao Inouye, Ryoma Kamikawa
Current Biology DOI: 10.1016/j.cub.2021.03.012本研究内容は朝日新聞デジタル版およびYahoo! ニュースでも紹介されています。
20年以上謎の生き物 正体判明 – Yahoo!ニュース環境DNA解析では、生物を培養することなく「どこに、どの生物が、どのくらいの量」存在するのかを知る手掛かりになります。しかしその一方で、DNA断片が未知の生物由来であった場合、生物の姿かたちや生態などの実態をDNA断片のみから読み取ることは困難なことが多いです。そのような謎の生物由来と考えられてきたDNAが20年以上前に報告されていました。未知の光合成生物である可能性は指摘されていましたが、実物がないので確証は得られませんでした。
本研究では上述した謎のDNAの持ち主と近縁である光合成性真核微生物を単離培養することに成功しました。本種はハプト藻類という光合成性真核生物のグループの新綱にあたると考えられる生物であったことから、謎のDNAはハプト藻類の新たな系統だったと考えることができました。本種は世界中の海に分布していることから、海洋の一次生産にとって無視できないレベルの貢献をしている可能性があります。
- 2021.03.22
-
本研究室 神川准教授の随想が京都大学広報誌に掲載されました。
以下のアドレスから閲覧することができます。
京大広報 2021.3 No.755
- 2021.02.22
-
2021年2月22日、歓送迎会および謝恩会を行いました。研究室を退職・卒業される方々、来年度から研究室に加わるメンバーも交えて、距離をとりながら歓談を楽しみました。
皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2021.1.12
-
2021.1.12当研究室で行われた研究がextremophiles誌に出版されました。
Diversity analysis of thermophilic hydrogenogenic carboxydotrophs by carbon monoxide dehydrogenase amplicon sequencing using new primers
Kimiho Omae,Tatsuki Oguro,Masao Inoue,Yuto Fukuyama,Takashi Yoshida & Yoshihiko Sako (2021),Extremophiles,
doi: 10.1007/s00792-020-01211-y水素生成型 CO 資化菌は環境中に発生する有毒な CO をエネルギー源となる水素に変換することで生態学的に重要な役割を担うと推測されています。しかし、環境中における存在量が低いことがボトルネックとなり、何がどこにいるかという生態学的知見は限定的でした。
本研究では水素生成と共役した CO 酸化を担う一部の系統の CO デヒドロゲナーゼを次世代シーケンサーによって高精度かつ高感度に検出する手法を新たに開発し、陸上温泉から未培養の新種を含む複数種の水素生成型 CO 資化菌を検出することに成功しました。本手法は有用なツールとして本菌の生態解明に寄与することが期待されます。
- 2020.12.25
-
本年度も当研究室OB・OGの皆様からお歳暮を頂きました。研究室一同でありがたく賞味させていただきました。
OB・OGの皆様、お心遣い頂きありがとうございました。
- 2020.11.30
-
2020年11月19~21日に開催された日本地球化学会第67回オンライン年会で本研究室の修士・博士課程の学生3名が口頭発表を行いました。また、特定研究員の井上真男さんがレビューセッションに参加しました。発表は大盛況で, M2の足立夕花さんが学生優秀賞を受賞しました。おめでとうございます!!題目
足立 夕花 始原的呼吸機構の遺伝学的解析に向けたParageobacillus thermoglucosidasiusにおける一酸化炭素酸化能欠失株の創出とその性状 武部 紘明 ミクロコズム培養系を用いた海洋性優占原核生物遷移過程におけるウイルス動態解析 富永 賢人 海洋原核生物も”密集”すればウイルスによる感染を受けるのか? 受賞コメント:このような賞をいただき非常に光栄です。卒業まで残り少しですが頑張ります。(足立)
- 2020.11.18
-
畜産資源学分野のD2佐藤さんが執筆された研究成果がPloSONEに掲載されました。
Calcium salts of long-chain fatty acids from linseed oil decrease methane production by altering the rumen microbiome in vitro
Yoshiaki Sato,Kento Tominaga,Hirotatsu Aoki,Masayuki Murayama,Kazato Oishi ,Hiroyuki Hirooka,Takashi Yoshida,Hajime Kumagai (2020),PLoS One, doi: 10.1371/journal.pone.0242158ルーメンへの 亜麻仁油(あまにゆ)添加によってメタン生成が抑制され、その細菌叢も変化することを示したものです。私たちにとっても大変興味深い共同研究となりました。何より佐藤さんおめでとうございます。
- 2020.11.09
-
11月9日に真如堂で撮影した記念写真です。紅葉がきれいに色づいており、和やかなひと時を過ごしました。


- 2020.11.04
-
光合成能を喪失した不等毛藻類の一種における葉緑体ゲノムおよびトランスクリプトームデータを解析した研究がFrontiers in Plant Scienceにアクセプトされました。(神川)
Highly reduced plastid genomes of the non-photosynthetic dictyochophyceans Pteridomonas spp. (Ochrophyta, SAR) are retained for tRNA-Glu-based organellar heme biosynthesis
Kayama M, Maciszewski K, Yabuki A, Miyashita H, Karnkowska A, Ryoma Kamikawa (2020),Front. Plant Sci., (in press) doi: 10.3389/fpls.2020.602455光合成能を喪失した藻類はほとんどの場合葉緑体を保持したままです。そして葉緑体ゲノムも保持されたままです。何故なんでしょう?
ということに部分的にではありますが答えることができたのがこの論文です。
おそらくアミノアシルtRNAを細胞質やミトコンドリアから非光合成性葉緑体に輸送できない限り、葉緑体ゲノムを喪失できないのではないか、と考えられます。
- 2020.10.20
-
本研究室の神川准教授が、公益財団法人農学会より第19回日本農学進歩賞を受賞しました。
「海洋真核微生物の次世代利用に向けたゲノム生物学的研究」
http://www.nougaku.jp/award/award1.2020.html学生時代から今日に至るまで、お世話になった皆様に感謝申し上げます。今後も、真核微生物の多様性解明と有効利用を目指し、さらに精進していく所存です。引き続きご指導ご鞭撻のほどよろしくお願い申し上げます。(神川)
- 2020.10.05
-
本研究室の神川准教授の研究が『みらいぶっくー学問・大学なび』に紹介されました。
https://www.miraibook-research.net/wakate/w2040/2020.09.17光合成能を喪失した緑藻類における葉緑体機能を解析した研究がBMC Biologyに出版されました。(神川)A non-photosynthetic green alga illuminates the reductive evolution of plastid electron transport systems.
Kayama M, Chen JF, Nakada T, Nishimura Y, Shikanai T, Azuma T, Miyashita H, Takaichi S, Kashiyama Y, Ryoma Kamikawa. (2020), BMC Biol. 18: 126ヒトや大腸菌のように光合成を行わない生物の多くは、基本的に葉緑体や光合成のための装置をもっていません。しかし、光合成を喪失するという進化を起こした藻類の多くは、非光合成性の葉緑体を保持しています。光合成をしなくなった藻類が葉緑体をもつ意義や、その背後にある分子機構を明らかにすることで、生物と光の関係を解き明かそうとしています。その一環として行った本研究では、光合成を喪失した後も、まだ電子伝達系の一部が機能し、様々な代謝の酸化還元バランスを取っていることが示唆されました。光合成をしない=(イコール)光合成装置のすべてが必要ない、ではないということです。
- 2020.09.17
-
真核生物における細胞内共生進化に関する短いレビューが出版されています。数か月前のものですが、紹介させていただきます。(神川)
Putative genome features of the relic green alga-derived nuclei in the dinoflagellates and the future perspectives as model organisms.
Nakayama T, Takahashi K, Ryoma Kamikawa, Iwataki M, Inagaki Y, Tanifuji G. (2020),Comm. Integr. Biol. 13:84-88
本年に入ってすぐに出版されたSarai et al. (2020) PNAS「Dinoflagellates with relic endosymbiont nuclei as novel models for elucidating organellogenesis」では真核微生物同士の細胞内共生による新たな葉緑体獲得現象の発見とその葉緑体が保持する縮退核を報告しました。今回の総説論文では、Sarai et al.での発見のどのような点が細胞内共生進化研究において重要であるかを概説するとともに、今後の研究の方向性を例示しています。
- 2020.09.16
-
修士課程の足立夕花さんによる論文がMicrobes and Environments誌に受理されました。足立さん、おめでとうございます!
Genetic Engineering of Carbon Monoxide-Dependent Hydrogen-Producing Machinery in Parageobacillus thermoglucosidasius
Yuka Adachi, Masao Inoue, Takashi Yoshida, Yoshihiko Sako (2020), Microbes Environ. (In press)
H2生成型CO酸化菌は有毒なCOガスをクリーンなH2に変換するため、H2生産型微生物触媒としての産業応用が期待できます。そこで私たちはParagebacillus thermoglucosidasiusに着目し、マーカーレス遺伝子欠失系を構築しました。その結果、本菌において今後の産業応用を目指すうえで基盤となるH2生成型CO酸化関連酵素群欠失株を作製することができ、本酵素群が本菌におけるH2生成型CO酸化に必須であることが明らかとなりました。
学部4回のときから行った初めての研究が原著論文として世に送り出され、本当に嬉しく思います。今回論文を書く過程で、「論理の幹」をつくり上げることがいかに大変であるかを理解し、実験系の組み立てや日頃からのディスカッションの積み重ねがとても重要であると学びました。先生方からの助言、研究チームや同期の励まし、さらに4回生の頃から指導をしてくれた井上さんの助けなしでは成し遂げられなかったです。どうもありがとうございました。
- 2020.08.07
-
大規模ブルームを形成するシアノバクテリアに感染するウイルスに絞って、そのゲノム特徴や増殖戦略、宿主とウイルスの共存機構をまとめた総説が受理されました。
Viruses of freshwater bloom‐forming cyanobacteria: Genomic features, infection strategies and coexistence with the host.
Daichi Morimoto, Sigitas Šulčius, Takashi Yoshida. (2020) Environ. Microbiol. Rep.
環境に大量に増殖するということは、ウイルスからの感染機会がぐんと上がるはずですが、ウイルスは宿主との均衡を保っています。不思議ですよね。そのウイルスと微生物の共存の仕組みをまとめていますので是非ご一読を。reportsの付いていない本体のからのレビュー投稿のお誘いだったのですが、対象範囲が狭くなったのが主因(だと思っています)で、 reportsの方に回ることになりました。今年は総説をまとめるのによい機会になっています!吉田
- 2020.07.31
-
吉田先生が責任著者の総説がAdvances in Applied Microbiology誌に受理されました。
Predetermined clockwork microbial worlds: Current understanding of aquatic microbial diel response from model systems to complex environments
Daichi Morimoto, Sigitas Šulčius, Kento Tominaga, Takashi Yoshida, Adv. Appl. Microbiol. In Press
水圏表層の低次生態系では、微細藻類の光合成により生じた日周リズムが、被食-捕食、ウイルス感染、共生といった様々な微生物間の相互作用を介して、多様な微生物群集全体に波及していくと考えられます。
本総説では、このような水圏低次生態系の日周期に注目し、分離培養系から明らかにされてきた日周期の分子メカニズムから、メタゲノム解析の発展により急速に理解が進みつつある環境微生物群集の日周動態まで、国内外の優れた研究成果を包括的に紹介し、本研究分野の今後の発展の一助となることを目指しました。
- 2020.07.31
-
特定研究員の井上真男さんの論文が Microbiology Resource Announcements 誌に受理されました。
Draft Genome Sequence of Thermanaeromonas sp. Strain C210 Isolated in the Presence of Carbon Monoxide
Masao Inoue, Ayumi Tanimura, Yuto Fukuyama, Suguru Okunishi, Hiroto Maeda, Takashi Yoshida, and Yoshihiko Sako(2020) Microbiol.Resour.Announc. 9:e00608-20.
鹿児島県山川湾海底堆積物から新規 Thermanaeromonas 属細菌の分離培養に成功し、ゲノム解析から本属細菌ゲノムの多様性と一酸化炭素代謝能を初めて明らかにしました。
- 2020.07.31
-
前回に続き当研究室OB・OGの皆様からお中元を頂きました。研究室一同、大変嬉しく思い、オンラインお茶会にてありがたく賞味いたしました。OB・OGの皆様、お心遣い頂きありがとうございました。
- 2020.07.15
-
本年度も当研究室OB・OGの皆様からお中元を頂きました。研究室一同、大変嬉しく思い、オンラインお茶会にてありがたく賞味いたしました。OB・OGの皆様、お心遣い頂きありがとうございました。
- 2020.05.22
-
博士課程の武部紘明さんの論文がMicrobes and Environments誌に受理されました。
Differential Responses of Coastal Prokaryotic Community to Phytoplanktonic Organic Matter Derived from Cellular Component and Exudate
Hiroaki Takebe, Kento Tominaga, Kentaro Fujiwara, Keigo Yamamoto and Takashi Yoshida (2020) Microbes and Environments, (In press)
海洋の原核生物は、植物プランクトンによって生成された有機物の多くを消費することから海洋物質循環に大きく貢献すると考えられており、その動態の理解が求められています。近年、環境中では原核生物の群集組成が日ごとに大きく変化することが報告されていますが、その機構の詳細は明らかではありませんでした。本論文では、世界的な海洋優占植物プランクトンであるHeterosigma akashiwo の有機物と海洋性原核生物群集を用いたミクロコズム培養系を構築し、植物プランクトンの生死と原核生物群集動態の関係性を調べました。その結果、植物プランクトンの死滅は周囲に生息する原核生物群集を短期間のうちに大きく変化させることが新たに明らかとなり、植物プランクトン-原核生物間の生態的関係の知見を大きく拡大しました。
- 2020.05.15
-
当研究室OBの森本大地さんの論文がMicrobiology Resource Announcements誌に受理されました。
Draft genome sequence of the astaxanthin-producing microalgae Haematococcus lacustris strain NIES-144
Daichi Morimoto, Takashi Yoshida, and Shigeki Sawayama. (2020) Microbiology Resource Announcements, (In press)
- 2020.04.28
-
特定研究員の井上真男さんの論文が Extremophiles 誌に受理されました。
Carbon Monoxide-dependent Transcriptional Changes in a Thermophilic,Carbon Monoxide-utilizing, Hydrogen-evolving Bacterium Calderihabitans maritimus KKC1 Revealed by Transcriptomic Analysis
Masao Inoue, Hikaru Izumihara, Yuto Fukuyama, Kimiho Omae, Takashi Yoshida and Yoshihiko Sako (2020) Extremophiles, (In press)
Calderihabitans maritimus KKC1 は鬼界カルデラ海底コアより分離された古代型の水素生成型一酸化炭素 (CO) 資化菌であり, 既知のバクテリアの中で最大である 7 つの CO 資化酵素を有しています。しかし, 本菌が CO シグナルに対してどのように応答するのかは分かっていませんでした。本論文では, 次世代シーケンサーを用いた網羅的転写変動解析 によって, CO 資化酵素遺伝子を含む本菌の CO 応答遺伝子群を多数明らかにするとともに, CO 資化に伴って変動する細胞内酸化還元状態を感知する新奇の転写応答機構を発見しました。
- 2020.04.13
-
2020年2月17日、歓送迎会および謝恩会を行いました。研究室を退職・卒業される方々、来年度から研究室に加わるメンバーも交えて、歓談を楽しみました。皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2020.04.01
-
博士課程の富永です。私が筆頭著者として投稿していた、海洋性Bacteroidetes門細菌に感染するウイルスに関する論文がFrontiers in Microbiology誌に受理されました。
In silico prediction of virus-host interactions for marine Bacteroidetes with the use of metagenome-assembled genomes
Kento Tominaga, Daichi Morimoto, Yosuke Nishimura, Hiroyuki Ogata and Takashi Yoshida (2020)Front. Microbiol.,(In press)
Bacteroidetes門は海洋で最も優占する従属栄養性細菌系統の一つで、特に植物プランクトンにより生産される有機物の分解者として生態学的に重要です。しかし、海洋性Bacteroidetes門細菌に感染するウイルスの分離例は一部の種に大きく偏っており、知見が不足していました。本研究では、海洋メタゲノムから報告されたBacteroidetes門細菌のゲノム(Metagenome assembled genomes, MAGs)と海洋ウイルスゲノムの比較により、細菌ゲノム上のウイルス感染の痕跡を探索し、81個の新規Bacteroidetes門細菌感染ウイルスを同定しました。これらの新規Bacteroidetes門細菌感染ウイルスには、水圏に広く分布するにも関わらず、宿主が明らかになっていなかったFar-T4系統ウイルスなど海洋の優占ウイルス系統が含まれました。
さらに、京都大学 化学研究所の緒方先生らの研究室を中心とした、巨大ウイルスの環境メタバーコーディング探索手法(メガプライマー法)に関する共同研究の成果が、Microorganisms誌に受理されました。緒方研究室の博士課程の同級生で、当研究室にも頻繁に来られて、一緒に実験をしてきたフロリアンさんの筆頭著者の論文です。
An optimized metabarcoding method for Mimiviridae
Florian Prodinger, Hisashi Endo, Yasuhiro Gotoh, Yanze Li, Daichi Morimoto, Kimiho Omae, Kento Tominaga, Romain Blanc-Mathieu, Yoshihito, Takano, Tetsuya Hayashi, Keizo Nagasaki, Takashi Yoshida, Hiroyuki Ogata (2020)Microorganisms,(In press)
- 2020.04.01
-
今年度より、神川龍馬先生が当研究室の准教授に着任されました。
また、今年から新たにメンバーとして加わる4回生4人との顔合わせを行いました。(ソーシャルディスタンス!!)4月1日付けにて、吉田天士先生の後任として海洋分子微生物学分野准教授に着任致しました神川龍馬と申します。これまで、嫌気環境に適応したミトコンドリアをもつ真核微生物や、光合成能を喪失した葉緑体をもつ微細藻類など、あまり教科書的にもなじみのない微生物を扱ってきました。これらの分離培養、ゲノム解析、遺伝子改変、生化学的解析を通じて、海洋真核微生物全体の進化・多様性と生態学的役割の解明、有用物質の生合成系の解明などに繋げていければと思っております。今後とも、ご指導ご鞭撻のほど、どうぞよろしくお願い申し上げます。
2020年4月1日 神川 龍馬


- 2020.02.12
-
博士課程の富永です。 私と吉田先生が共著者として参加させていただいた国際共同研究が、Environmental microbiology reportsに受理されました。 スペイン アリカンテ大学のグループを中心とした、全海洋で最も優占する37−F6ウイルスの日周期の研究です。
Diel cycling of the cosmopolitan abundant Pelagibacter virus 37‐F6: one of the most abundant viruses on earth
Francisco Martinez‐Hernandez, Elaine Luo, Kento Tominaga, Hiroyuki Ogata, Takashi Yoshida, Edward F. DeLong, Manuel Martinez‐Garcia. (2020). Environ Microbiol Rep,doi.org/10.1111/1758-2229.12825
- 2019.12.06
-
2019年9~11月は当研究室より多くの方が学会に参加し、発表を行いました。発表を行った皆様、お疲れ様でした!!
2019年11月29,30日、第12回目となる「藍藻の分子生物学2019」が、かずさDNA研究所近くの「かずさアカデミアホール」で開催され、当研究室の吉田先生が招待講演を行い、博士後期課程の富永さんがポスター発表を行いました。 共同研究に繋がる出会いも多く、将来に繋がるお土産の多い会となりました。
招待講演吉田 天士 Cross-omics解析から明らかになった海洋シアノファージの感染動態 ポスター発表
富永 賢人 長期観測による海洋シアノバクテリア-ウイルス相互作用の解明 2019年11月23,24日、北里大学において行われた国際水圏メタゲノムシンポジウムで特定研究員の井上真男さんが口頭発表を行いました。
題目井上 真男 Metagenomic study for hydrogenogenic carbon-monoxide oxidizers as rare biosphere 2019年11月23日、近畿大学農学部において日本水産増殖学会第18回大会・令和元年度日本水産学会近畿支部例会が開催されました。 M2の武部さん、中川さん、野澤さんが口頭発表を行い、野澤さんが優秀発表賞を受賞しました。 野澤さん、おめでとうございます!!
題目武部 紘明 海洋性微細藻類由来有機物が原核生物群集に与える影響に関する研究 中川 哲 アオコ原因ラン藻 Microcystis aeruginosa のウイルス感染細胞直接定量法の確立 野澤 理香 海底堆積物を用いた集積培養系における一酸化炭素酸化菌の動態解析 受賞コメント: この度はこのような賞をいただき大変光栄に存じます。日頃よりご指導いただきました研究室の皆様、ご協力いただいた共同研究者の皆様、そして本学会を運営してくださった皆様に心より御礼申し上げます。今後とも一層精進してまいりますので、どうぞよろしくお願いいたします。
2019年11月22日、京都大学大学院農学研究科に開設された『糸状菌・環境インターフェイス工学講座』の開設記念式典・シンポジウム『IFOがつなぐ京大微生物学のフロントライン』が開催され、当研究室の吉田先生による研究分野の紹介が行われ、研究室の学生5名もポスター発表を行いました。 京都大学の様々な学部から、各分野をリードする微生物系研究者が一堂に会する非常に貴重な機会でした。同じ大学内にこんな研究をしている人がいたのか、という新たな発見もあり、ポスター議論も盛り上がりました。
題目
日野 太貴 新規 Parageobacillus 属細菌の生理・ゲノム特性による一酸化炭素酸化能の多様性に関する研究 足立 夕花 Parageobacillus thermoglucosidasius における水素生成型一酸化炭素酸化関連酵素群の機能解明に向けた遺伝子組換え研究 西田 志穂 鹿児島県山川湾沖海底堆積物における一酸化炭素が促進する微生物群集の評価 藤原 健太郎 16S-23S rDNA ITS 領域を標的とした海洋優占系統 SAR86 細菌の高解像度動態解析 富永 賢人 大阪湾での経時的動態観測で探る海洋微生物-ウイルス群集相互作用ネットワーク 2019年11月16,17日、京都大学桂キャンパスで行われた第20回極限環境生物学会年会において特定研究員の井上真男さんが口頭発表を, M1の足立夕花さんと岡元俊輔さんがポスター発表を行いました。発表は大盛況で, M1の足立夕花さんが優秀ポスター賞を受賞しました。おめでとうございます !!
題目
井上 真男 Parageobacillus 属細菌における一酸化炭素利用能の多様性 足立 夕花 Parageobacillus thermoglucosidasius における水素生成型一酸化炭素酸化関連酵素群の機能解明に向けた遺伝子組換え研究 岡元 俊輔 Parageobacillus 属細菌における水素取り込み型ヒドロゲナーゼを介したエネルギー獲得機構に関する研究 2019年9月10~13日に山梨大学 甲府キャンパスで開催された日本微生物生態学会第33回大会で本研究室の修士・博士課程の学生4名が口頭発表を行いました。
題目
日野 太貴 水素生成一酸化炭素酸化細菌Parageobacillus sp.G301株の生理性状およびゲノム解析 武部 紘明 海洋性真核微細藻類が原核生物群集構造に与える影響に関する研究 富永 賢人 未分離の海洋性Bacteroidetes門細菌感染ウイルスの生物情報学的探索 礒崎 達大 シアノウイルス-宿主相互作用解明に向けた海洋シアノバクテリアの高純度分手法の確立 2019年9月2~6日、九州大学において行われた第15回国際好熱菌学会で特定研究員の井上真男さんが口頭発表を行いました。加えて、特定研究員の福山宥斗さんとM1の岡元俊輔さんがポスター発表を行いました。
題目
井上 真男 Redox-balancing in Carbon Monoxide-utilization of a Thermophilic, Hydrogenogenic Carboxydotroph Calderihabitans maritimus KKC1 Revealed by a Comparative Transcriptomic Study 福山 宥斗 Insight into Hydrogenogenic Carbon Monoxide Metabolism in Carboxydothermus pertinax Revealed by Comparative Genomic Analysis, Growth Experiments and Whole Transcriptome Analysis 岡元 俊輔 Isolation, Growth Profile, and Genome Sequence of Parageobacillus thermoglucosidasius Strain TG4, a Thermophilic, Hydrogenogenic, Carboxydotrophic Bacterium, First Isolated from a Marine Sediment
- 2019.11.15
-
2019年11月4日、京都大学化学研究所において2019年 環境ウイルス研究集会が開催されました。 当研究室OBの森本さんと博士課程の富永さんが口頭発表を行い、森本さんが最優秀発表賞、富永さんが優秀発表賞を受賞されました。 森本さん、富永さん、おめでとうございます!!
研究集会HP http://www.microbial-ecology.jp/?p=8672題目
森本大地 アオコ原因ラン藻Microcystis aeruginosaと相互作用する新規ウイルスの環境動態解析 受賞コメント: この度はとても光栄な賞をいただき大変嬉しく思っております。 今回はアオコ原因ラン藻Microcystis aeruginosaに感染する未分離ウイルスをメタゲノム解析により明らかにし、 環境中におけるウイルス転写動態と宿主遺伝子の応答をメタトランスクリプトーム解析により調べた研究 (Morimoto et al.,Appl. Environ. Microbiol., 2019) を中心にご紹介させていただきました。 この最優秀講演賞は先輩方の素晴らしい研究成果と周囲の皆様のサポートの積み重ねで得られた成果だと考えています。 この賞を励みに、これからも初心を忘れずにより素晴らしい研究成果を挙げられるよう邁進していきたいと思います。 今後ともよろしくお願い致します。
題目
富永賢人 大阪湾での経時的動態観測で探る海洋微生物-ウイルス群集相互作用 受賞コメント:今回の受賞は、皆様からの叱咤激励の証と思いますので、なお一層気を引き締めて研究を進めていきたいと思います。改めて、日頃支えてくださっている皆様に感謝を申し上げます。本発表の内容をさらに磨きをかけ、国際的な成果として論文発表できるよう、邁進いたします!


- 2019.11.14
-
2019年10月25日、海洋深層水中の微生物・ウイルス動態調査を目的に、高知県室戸市の高知県海洋深層水研究所においてサンプリングを行いました。同研究所は日本で初めて設置された海洋深層水研究施設で、約2.6kmにおよぶ取水管により、湧昇により昇ってきた深層水を水深約320 mから常時汲み上げています。湧昇域は全世界の海域の0.1%しか存在せず、このような海洋深層水を常時採水できる施設は非常に希少で、国際的にも新規な知見が期待されます。研究所の皆様のご協力のもと、なんとか天候にも恵まれて、貴重なサンプルを採取できました。



- 2019.09.16
-
当研究室のOBである望月智弘先生が、Themophile2019への招待講演で来日されたMart Krupovic博士と共に研究室へと足を運んで下さいました。お話を聞かせて頂き、ありがとうございました。今後ともよろしくお願いいたします。

- 2019.09.16
-
今月公表された「Co-occurrence of broad and narrow host-range viruses infecting the toxic bloom-forming cyanobacterium Microcystis aeruginosa. 」の掲載号であるApplied and Environmental Microbiology vol. 85 no. 18において、研究フィールドである広沢池のアオコの写真が表紙を飾りました。

- 2019.08.08
-
本年度も当研究室OB・OGの皆様からお中元を頂きました。研究室一同、大変嬉しく思い、ありがたく賞味いたしました。OB・OGの皆様、お心遣い頂きありがとうございました。

- 2019.07.25
-
博士課程卒業生の森本大地さん(現 京都大学大学院 農学研究科 海洋環境微生物学研究室 特定助教)による論文が、Applied and Environmental Microbiologyに受理されました。森本さん、おめでとうございます!
Co-occurrence of broad and narrow host-range viruses infecting the toxic bloom-forming cyanobacterium Microcystis aeruginosa.
Morimoto, D., Tominaga, K., Nishimura, Y., Yoshida, N., Kimura, S., Sako, Y., Yoshida, T. (2019) Appl. Environ. Microbiol. (In press)
ラン藻ミクロキスティスは肝臓毒生産能を有し、世界中の湖沼で異常増殖してアオコを形成するため、重要な生物種と捉えられています。本種は極めて多くのウイルス耐性遺伝子を有し、多種多様なウイルスと環境中で相互作用すると示唆されてきましたが、本種感染ウイルス分離例は当研究室保有のMa-LMM01に限られていました。本研究では京都府広沢池にてウイルスメタゲノム解析をおこない、24時間に渡ってミクロキスティスとウイルスの包括的転写解析をおこないました。その結果、大きく3つのグループから成る15の新規本種感染ウイルスゲノムを明らかにしました。また環境中にはMa-LMM01のような非常に限られたミクロキスティス株にのみ感染可能な狭域宿主ウイルスと、様々なミクロキスティス株に感染可能な広域宿主ウイルスが共存することを見出しました。包括的転写解析より、ミクロキスティスは特に後者による感染に応じてウイルス耐性遺伝子を発現させることが示唆されました。また本成果に関わる写真がApplied and Environmental Microbiology vol. 85 no. 18の表紙を飾る予定です。
- 2019.07.25
-
特定研究員の福山宥斗さんによる論文が、Microbiology Resource Announcementsに受理されました。福山さん、おめでとうございます!
Draft Genome Sequences of Two Thermophilic Bacteria Moorella spp., Isolated from an Acidic Hot Spring in Japan
Fukuyama, Y., Tanimura, A., Inoue, M., Omae, K., Yoshida, T., Sako, Y. (2019) Microbiol. Resour. Announc., DOI:10.1128/MRA.00663-19
水素生成型CO資化性菌は有毒なCOを酸化し、利用しやすい水素に変換するため 微生物生態学や応用利用面で注目されていますが、その分離例は少なく、ゲノム解析例は さらに限られています。本論文では私達がメインフィールドとする鹿児島県鰻温泉より 分離した新規株のゲノム解析を行いました。その結果、本分離株が既知のMoorella属細菌と系統的に異なる 新種に相当すること、またCO酸化と水素生成が可能なゲノム特性を有することを見出しました。 本成果により、水素生成型CO資化性菌のゲノムデータの拡充に成功しました。
- 2019.05.08
-
博士課程の大前公保さんによる論文が、Archives of microbiologyに受理されました。大前さん、おめでとうございます!
Diversity and distribution of thermophilic hydrogenogenic carboxydotrophs revealed by microbial community analysis in sediments from multiple hydrothermal environments in Japan
Omae, K., Fukuyama, Y., Yasuda, H., Mise, K., Yoshida, T., Sako, Y. (2019) Arch. Microbiol., doi.org/10.1007/s00203-019-01661-9
- 2019.04.30
-
2019年4月19日、本年度は新たに11人の新メンバーを迎え歓迎会を行いました。談笑に花を咲かせ、楽しい時間を過ごしました。一年間、共に頑張っていきましょう。

- 2019.04.12
-
特定研究員の福山宥斗さんによる論文が、Extremophilesに受理されました。福山さん、おめでとうございます!
Transcriptome analysis of a thermophilic and hydrogenogenic carboxydotroph Carboxydothermus pertinax
Fukuyama, Y., Omae, K., Yoshida, T., Sako, Y. Extremophiles., 23, 389-398(2019)
- 2019.04.04
-
新年度の顔合わせを行いました。今年は新たに4回生4人、修士6人、PD研究員1人を迎えました。これで4回生4人、修士課程11人、博士課程3人、PD研究員2人となりました。メンバーが大幅に変わりましたが、今年度も賑やかな研究室になりました。これからよろしくお願いします。

- 2019.04.01
-
4月1日付けにて、左子芳彦先生の後任として海洋分子微生物学分野教授を拝命致しました吉田天士でございます。これまで、シアノバクテリアを含む微細藻類の生理・生態学的研究を行ってきました。この過程でこうした微生物に感染するウイルスの面白さにひかれ、新たにゲノム科学的手法による微生物ウイルス解析にも取り組むようになりました。今後はウイルス-微生物相互作用を組み込んだ包括的な海洋微生物生態学分野を確立していきます。もちろん、左子先生が確立されてきた熱水環境における新規難培養性微生物の分離培養技術も継承し、新たな遺伝子資源探索も進めてまいります。是非とも、ご支援いただければ幸甚でございます。 5月半ばをめどにホームページを改訂いたします。まずは、ご挨拶申し上げます。
2019年4月1日 吉田 天士


- 2019.03.30
-
2019年3月26~29日、東京海洋大学において平成31年度水産学会春季大会が開催され、左子先生が日本水産学会賞を受賞されました。左子先生おめでとうございます。当研究室から6名が口頭発表を行いました。会場では多くの質問やコメントを頂き、大変有意義な学会発表となりました。
題目磯崎 達大 セルソータ―による高純度細胞分取法を利用した海洋ラン藻の細胞内ウイルス定量法の確立の試み 森本 大地 アオコ原因ラン藻Microcystis aeruginosaと相互作用する新規ウイルスの定量的検出法の確立と環境動態解析 大前 公保 全原核生物の 16S rRNA 系統における一酸化炭素 (CO) デヒドロゲナーゼの分布パターンを利用した水素生成型 CO 資化菌の分子生態 富永 賢人 大阪湾の海洋微生物・ウイルスの生態学的相互作用解析 福山 宥斗 クロスオミックス解析を用いた水素生成型一酸化炭素 (CO) 資化性菌のCO代謝の解明 井上 真男 火山帯水圏環境由来 Parageobacillus 属水素生成型一酸化炭素 (CO) 資化菌のゲノム解析 
- 2019.03.25
-
2019年3月12日、招へい研究員としてリトアニアより来日されていたSigitas Sulcius博士による最終講義、「バルト海におけるウイルス生態学」が行われました。講義の後には、縄田農学研究科長より、本交流プログラムの修了証書が手渡されました。Sigitasさん、1年間大変お疲れ様でした。


- 2019.03.25
-
2019年3月9日、今年度をもって退職される、左子芳彦教授の最終講義および退職記念祝賀会が開催されました。微生物の研究をはじめ、さまざまな分野で活躍される研究室OB・OG、関係者の方々が一堂に会し、左子先生のご退職をお祝いしました。大変賑やかで温かく、思い出に残る1日となりました。



- 2019.02.20
-
2019年2月15日、歓送迎会および謝恩会を行いました。来年度から研究室に加わる方々も交え、歓談を楽しみ、研究室を卒業される方々の門出をお祝いしました。皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2019.01.21
-
特定研究員の井上真男さんによる論文2報が、Frontiers in MicrobiologyおよびMicrobiology Resource Announcementsにそれぞれ受理されました。井上さん、おめでとうございます!
Draft genome sequence of Parageobacillus thermoglucosidasius strain TG4, a hydrogenogenic carboxydotrophic bacterium isolated from a marine sediment
Inoue, M., Tanimura, A., Ogami, Y., Hino, T., Okunishi, S., Maeda, H., Yoshida, T., Sako, Y. (2019) Microbiol. Resour. Announc., 8:e01666-18
Structural and Phylogenetic Diversity of Anaerobic Carbon-monoxide Dehydrogenases
Inoue, M., Nakamoto, I., Omae, K., Oguro, T., Ogata H., Yoshida, T., Sako, Y. (2019) Front. Microbiol., 9:3353. doi: 10.3389/fmicb.2018.03353
- 2018.12.27
-
2018年12月21日、大掃除と忘年会を行いました。今年も一年間おつかれさまでした。来年もさらに楽しく研究を発展させることができるよう、皆で力を合わせて頑張りましょう。

- 2018.12.12
-
2018年12月8日、滋賀県立琵琶湖博物館において日本水産学会近畿支部 平成30年度後期例会が開催されました。 当研究室から博士課程の磯崎達大さんが口頭発表を行い、優秀発表賞を受賞されました。 磯崎さん、おめでとうございます!!
題目磯崎 達大 海洋における原核微生物とウイルスの感染ペア同定に向けた特定細胞集団の高純度回収法 
- 2018.12.05
-
2018年11月27~29日、奈良県東大寺総合文化センターにおいてThe 5th International Conference of Members of the Genus Flavobacterium ‘Flavobacterium 2018’ が開催され、当研究室から博士課程の富永さんが口頭発表を行いました。国内外の魚類病原性フラボバクテリアの専門家の方々から、我々のオミクス解析による微生物-ウイルス生態学研究手法について多数のご意見を頂き、非常に有意義な会となりました。
題目富永 賢人 New lineages of marine Bacteroidetes viruses identified by MAGbased host prediction and their ecological interaction with hosts bacteria
- 2018.11.19
-
2018年10月27日、京都大学理学部において環境ウイルス集会が開催され、本研究室のリトアニアからの招へい研究員Sigitas Sulcius博士、木村さん、森本さん、富永さんがそれぞれ口頭発表を行いました。多様なウイルス研究者の方々のお話を聞くことができ、今後の研究にむけて刺激を受けた集会となりました。
題目Sigitas Sulcius Cyanophage effect on cyanotoxins dynamics 
木村成子 Natural population dynamics of Microcystis aeruginosa associated with its infectious phages detected by the leader-end CRISPR fragment analysis 森本大地 Cross-Omics analysis of a toxic bloom-forming cyanobacterium Microcystis aeruginosa and its viruses 富永 賢人 Population dynamics of marine microbes and viruses and their ecological interaction in Osaka bay
- 2018.11.13
-
2018年10月18日、応用生物科学専攻において博士論文申請講演会が行われ、 当研究室から森本大地さんと福山宥斗さんが口頭発表を行いました。
題目森本大地 Comprehensive studies on transcriptional dynamics of cyanoviruses infecting a bloom-forming cyanobacterium Microcystis aeruginosa 福山宥斗 Genomic and transcriptional studies on hydrogenogenic carboxydotrophic bacteria
- 2018.09.25
-
2018年9月15~18日、広島大学において平成30年度水産学会秋季大会が開催され、当研究室から院生5名が口頭発表を行いました。会場では多くの質問やコメントを頂き、大変有意義な学会発表となりました。
題目富永 賢人 海洋微生物・ウイルス間相互作用の解明に向けた海洋真核微生物の群集ダイナミクス解析 中元 一星 一酸化炭素 (CO) 資化性菌の鍵酵素、COデヒドロゲナーゼの系統学的多様性 武部 紘明 海洋性真核微細藻類が原核生物群集構造に与える影響に関する研究 和泉原 光 網羅的転写解析による海洋性一酸化炭素 (CO) 資化性菌のCO代謝経路の探索 日野 太貴 鹿児島湾海底堆積物からの水素生成型一酸化炭素 (CO) 資化性菌の分離と性状解析 
- 2018.09.25
-
2018年8月26日から9月11日にかけて日本学術振興会二国間交流事業共同研究で、当研究室から吉田先生とSigitasさん、院生2名がリトアニアを訪問しました。
前半ではNational Recerch Centre (NRC) やヴィリニュス大学の共同研究者の方々とミーティングやセミナーを行い、後半ではバルト海に隣接するCuronian Lagoonにおいてラン藻ブルームサンプルを採取しました。
国際的に活躍されている研究者の方々との交流を通し多くの刺激を受け、有意義な時間を過ごすことができました。




- 2018.08.08
-
2018年8月3日、鹿児島湾において海水サンプルおよびコアサンプルの採取を行いました。昨年に引き続き、鹿児島大学水産学部の前田広人教授のご尽力のもと、南星丸に乗船させていただきサンプリングを行いました。 船員の方々をはじめ、関係者の皆様の多大なご協力により貴重なサンプルを採取することができました。今後の成果が期待されます。

- 2018.06.28
-
2018年6月17~21日、アメリカのネブラスカ州においてAquatic Virus Workshop 9が開催され、当研究室からSigitasさん、木村さん、森本さんがそれぞれ口頭発表を行いました。 また、会場で参加者のみなさんと記念写真を撮影しました。
題目木村成子 Microcystis population dynamics associated with its infectios phages detected by the leader-end CRISPR fragment analysis in a natural pond 森本大地 Transcriptome analysis of a toxic bloom-forming cyanobacterium Microcystis aeruginosa during Ma-LMM01 phage infection Sulcius Sigitas Baltic sea cyanopages: diversity, distribution and insights into virus-host interaction 
- 2018.05.17
-
博士課程の福山宥斗さんによる論文がApplied and Environmental Microbiologyに受理されました。福山さんおめでとうございます!!
Insight into Energy Conservation via Alternative Carbon Monoxide Metabolism in Carboxydothermus pertinax Revealed by Comparative Genome Analysis.
Yuto Fukuyama, Kimiho Omae, Yasuko Yoneda, Takashi Yoshida, Yoshihiko Sako. (2018). Appl Environ Microbiol,84,e00458-18
- 2018.05.01
-
2018年4月23日、本年度は新たに10人の新メンバーを迎え歓迎会を行いました。ピアノ、歌、ダンス等皆で大変盛り上がり楽しい時間を過ごしました。一年間、共に頑張っていきましょう。

- 2018.05.01
-
当研究室OBの本田貴史さん(現 公益財団法人神戸医療産業都市推進機構)と院生の森本大地さんによる論文がMarine Biotechnologyに受理されました。本田さん、森本さんおめでとうございます!!
LexA binds to transcription regulatory site of cell division gene ftsZ in toxic cyanobacterium Microcystis aeruginosa.
Takashi Honda., Daichi Morimoto., Yoshihiko Sako and Takashi Yoshida.
- 2018.04.19
-
2018年4月14日~16日に高知県の土佐ロイヤルホテルにおいて第四回新学術領域「ネオウイルス学」領域班会議が開催され、当研究室から博士課程の富永さんが参加し、ポスター発表を行いました。
ポスター発表 題目富永賢人 大阪湾湾口部における未分離の海洋性Bacteroidetes門細菌およびウイルスの時系列変動の解析
- 2018.04.09
-
2018年4月5日、本研究室の名誉教授 石田祐三郎先生が培根会に出席された後、来研されました。今年85歳になられた先生は大変お元気で、昔の懐かしいお話をされて記念写真を撮り、楽しい時間を過ごしました。

- 2018.04.09
-
2018年3月27日、東京海洋大学(品川キャンパス)において、平成30年度日本水産学会大会春季大会が開催され、当研究室から4つの口頭発表が行われました。
口頭発表 題目
井上真男 一酸化炭素 (CO) 資化性菌の鍵酵素, CO デヒドロゲナーゼにおける構造と機能の多様性 大前公保 火山性環境における一酸化炭素資性菌の分布および多様性の解明 森本大地 アオコ原因ラン藻Microcystis aeruginosaの新規ウイルスの網羅的探索 富永賢人 未分離の海洋性バクテロイデス門細菌感染ウイルスゲノムの探索と同定
- 2018.04.09
-
ポスドクの木村成子さんの論文がFrontiers in Microbiologyに掲載されました。木村さんおめでとうございます!!
Incomplete selective sweeps of Microcystis population detected by the leader-end CRISPR fragment analysis in a natural pond.
Kimura, S., Uehara, M., Morimoto, D., Yamanaka, M., Sako, Y., Yoshida, T. (2018). Front. Microbiol. , fmicb.2018.00425
- 2018.04.09
-
2018年3月5日~7日、京都大学 桂キャンパス 船井哲良記念講堂において「第12回日本ゲノム微生物学会年会」が開催され、7日のシンポジウムにおいて吉田先生が講演を行いました。
講演題目吉田天士 The cross-omics of marine viruses: ecological perspective
- 2018.03.01
-
2018年2月20日、歓送迎会・謝恩会を行いました。就職される修士修了5人、建築を勉強される4回生1人、来年度から新たにラボに加わる方々も交え、歓談を楽しみました。皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2018.02.16
-
2018年2月6日、修士論文発表会が行われました。当研究室から5名が口頭発表を行いました。
口頭発表 題目笈西優 新規好気性超好熱古細菌の分離および生理性状・ゲノム特性に関する研究 大黒達希 火山帯環境における好熱性一酸化炭素資化性菌の多様性解析 佐々木彩 Droplet Digital PCR法を用いた直接定量法による有毒ラン藻Microcystis aeruginosaとその感染性ウイルス検出法の確立 晴気七菜 高解像度16S rRNAメタゲノム解析を用いた大阪湾湾口部における原核生物の群集ダイナミクス 山中ももこ ウイルス感染履歴CRISPR配列に基づくMicrocystisの個体群ダイナミクス 2018年2月13日、課題研究発表会が行われました。当研究室から3名がポスター発表を行いました。
口頭発表 題目鈴木友也 高知県浦ノ内湾における藻類ブルームに応答する海洋性原核生物の時系列変動解析 武部紘明 海洋性真核微細藻類による原核生物群集に与える影響に関する研究 日野太貴 海底堆積物コア中の古代型一酸化炭素資化性菌の探索に向けた芽胞精製法の確立
- 2018.02.02
-
吉田天士先生および院生の森本大地さんの論文がそれぞれThe ISME JournalおよびFrontiers in Microbiologyに掲載されました。吉田先生、森本さんおめでとうございます!!
Locality and diel cycling of viral production revealed by a 24 h time course cross-omics analysis in a coastal region of Japan.
Yoshida, T., Nishimura, Y., Watai, H., Haruki, N., Morimoto, D., Kaneko, H., Honda, T., Yamamoto, K., Hingamp, P., Sako, Y., Goto, S., Ogata, H. (2018). The ISME Journal , s41396-018-0052-x
Transcriptome Analysis of a Bloom-Forming Cyanobacterium Microcystis aeruginosa during Ma-LMM01 Phage Infection.Morimoto, D., Kimura, S., Sako, Y., Yoshida, T. (2018). Front. Microbiol. 9, fmicb.2018.00002
- 2017.12.21
-
2017年12月21日、忘年会を行いました。今年一年おつかれさまでした。2018年はさらなる成果をあげるため一丸となって研究を進めていきましょう!

- 2017.12.21
-
2017年12月10日、京都大学農学部において日本水産学会近畿支部後期例会が開催され、当研究室から修士課程の佐々木彩さんが口頭発表を行い、優秀発表賞を受賞しました。おめでとうございます。
口頭発表 題目佐々木彩 Droplet Digital PCR法を用いた有毒ラン藻Microcystis aeruginosaの直接定量法の確立
- 2017.11.13
-
2017年9月22~24日に東京海洋大学で開催された日本水産学会創立85周年記念国際シンポジウムで、修士課程の晴気七菜さんおよび大神優祐さんがポスター賞を受賞されました。おめでとうございます。
ポスター発表 題目
晴気七菜 Seasonal dynamics of eukaryotic microbes and prokaryotes in Osaka Bay, Japan 大神優祐 Isolation and characterization of a new carboxydotrophic bacterium
- 2017.11.13
-
2017年10月29日~31日に静岡県のホテルラフォーレ修善寺において第三回新学術領域「ネオウイルス学」領域班会議が開催され、当研究室から博士課程の富永さんが参加し、ポスター発表を行いました。
ポスター発表 題目富永賢人 ウイルス分類ツールViPTreeによる大阪湾における海洋ウイルス群集の季節変動の解析
- 2017.10.05
-
2017年9月24日に、鹿児島大学水産学部の前田広人教授のご尽力により南星丸に乗船させていただきサンプリングを行いました。鹿児島湾奥の若尊カルデラ近傍において、海水サンプルおよびコアサンプルの採取を行いました。今回は船員の方々をはじめ、関係者の皆様の多大なご協力により貴重なサンプルを採取することができました。成果が期待されます。

- 2017.10.05
-
9月18日に来日され、二週間交流事業で研究・交流を行ったSigitas Sulcius博士がリトアニアに帰国することになりました。Sigitasさんとは研究についてだけではなく、両国の文化などについての様々なお話ができ、とても有意義な時間を過ごすことができました。最後に、Sigitasさん、スタッフおよび学生で記念写真を撮りました。

- 2017.10.05
-
2017年9月22~24日に東京海洋大学品川キャンパスにおいて日本水産学会創立85周年記念国際シンポジウムが開催されました。当研究室からは左子先生、吉田先生、院生5名が参加し、口頭発表またはポスター発表が行われました。またリトアニアのNature Research CentreのSigitas Sulcius博士による招待講演も行われました。
口頭発表 題目左子芳彦 Comprehensive, spatiotemporal study and applied research of carboxydotrophs 吉田天士 What does viral metagenomics provide for future marine ecology and resources 森本大地 Transcriptome analysis of Mycrocystis aeruginosa during Ma-LMM01 phage infection 大前公保 Genomic analysis of Calderihabitans maritimus KKC1, a thermophilic hydrogenogenic carboxydotrophic bacterium isolated from marine sediment ポスター発表 題目
笈西優 Isolation and characterization of a novel aerobic and hyperthermophilic archaeon isolated from a hot spring in Japan 晴気七菜 Seasonal dynamics of eukaryotic microbes and prokaryotes in Osaka Bay, Japan 大神優祐 Isolation and characterization of a new carboxydotrophic bacterium 講演 題目
Sigitas Sulcius Toward better understanding of the Baltic Sea ecosystem services through the studies of cyanobacteria-virus interactions 
- 2017.10.05
-
8月29日から9月2日にかけて日本学術振興会二国間交流事業共同研究で、当研究室から吉田先生と院生2名がリトアニアを訪問しました。リトアニアではNational Research Centre (NRC) やヴィリニュス大学において共同の研究者の方々とミーティングやセミナーを行いました。研究だけではなく両国の文化についても理解を深めることができ、非常に貴重な体験となりました。

- 2017.08.10
-
トカラ列島鬼界カルデラの海底コアから分離された、新属種の一酸化炭素資化性好熱菌の ゲノム解析に関する論文がAEM8月号に掲載され、その新奇性や特異性から大いに注目を集めて本種の 凝集した電顕写真が表紙に掲載され、記念すべき論文となりました。
Genomic analysis of Calderihabitans maritimus KKC1, a thermophilic hydrogenogenic carboxydotrophic bacterium isolated from marine sediment.
Omae, K., Yoneda, Y., Fukuyama, Y., Yoshida, T., and Sako, Y. (2017). Appl Environ Microbiol . 83(15) e00832-17 doi:10.1128/AEM.00832-17

- 2017.06.06
-
2017年6月3日、4日に東北大学(青葉山新キャンパス)において、第19回マリンバイオテクノロジー学会仙台大会が開催されました。当研究室からは2つの口頭発表が行われました。
口頭発表 題目福山宥斗 オーミックス解析を用いた水素生成型一酸化炭素 (CO) 資化性菌の CO 代謝とその制御系の解明 大前公保 水素生成型一酸化炭素資化性菌の生態学的機能解明に向けた陸上熱水環境における微生物群集構造解析
- 2017.06.06
-
2017年5月29日、本年度は新たに10人の新メンバーを迎え歓迎会を行いました。バイオリン、ピアノ、歌、ダンス等皆で大変盛り上がり楽しい時間を過ごしました。一年間、共に頑張っていきましょう。

- 2017.06.05
-
4月17日に、最近息子さんが誕生し先端医療のお仕事を始めたOBの本田貴史さんとご家族が来研され、歓談後記念写真を撮りました。お子さんの健やかな成長をお祈りいたします。

- 2017.04.17
-
新年度の顔合わせを行いました。今年は新たに4回生4人、修士4人、博士1人、PD研究員3人を迎えました。これで4回生4人、修士課程9人、博士課程5人、PD研究員3人となりました。メンバーが大幅に変わりましたが、今年度も賑やかな研究室になりました。これからよろしくお願いします。

- 2017.04.04
-
2017年3月26~30日、東京海洋大学(品川キャンパス)において、平成29年度 日本水産学会大会春季大会が開催され、当研究室から3つの口頭発表が行われました。
口頭発表 題目福山宥斗 網羅的転写解析による一酸化炭素 (CO) 資化性菌のCO代謝制御系の解明 森本大地 アオコ原因ラン藻Microcystis aeruginosaにおけるファージ感染時の網羅的転写動態解析 大前公保 陸上熱水環境における微生物群集構造解析および一酸化炭素資化性菌探索の試み また学会期間中のお昼に、当研究室OBでダウ・ケミカル日本(株)で活躍されている高尾郷介さんと歓談し記念写真を撮影しました。

- 2017.02.21
-
2017年2月17日、来年度よりラボを移動される研究員1人、就職される修士修了3人、理学研究科へ進学される4回生1人の5人の送別会を行いました。来年度から新たにラボに加わる方々も交え、歓談を楽しみました。皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

2017年2月13日、課題研究発表会が行われました。当研究室から4名がポスター発表を行いました。
ポスター発表 題目和泉原光 生理学的研究に向けた海洋性一酸化炭素資化性菌培養系の確立 大神優祐 新規一酸化炭素資化性菌の分離の試み 澤田佳奈 大阪湾における原核生物の季節的な動態解析 福田将矢 Microcystis aeruginosaの遺伝的変異に起因する表現型の変異に関する研究 2017年2月15日、修士論文発表会が行われました。当研究室から3名が口頭発表を行いました。
口頭発表 題目中田舞 ファージとの相互作用によるMicrocystis aeruginosaの遺伝的変異に関する研究 弓矢真穂 Isolation and characterization of a novel temperate virus Aeropyrum globular virus 1 infecting hyperthermophilic archaeon Aeropyrum (超好熱古細菌に感染する新規溶原化ウイルスAeropyrum globular virus 1の分離と性状解析) 吉安勇人 大阪湾湾口における18S アンプリコン解析を用いた真核微生物群集の季節的変動
- 2017.02.02
-
2016年12月13~15日、CO資化性菌の分離とメタゲノム解析を目的に、鹿児島県霧島市の山乃城温泉および同県指宿市の鰻温泉においてサンプリングを行いました。約1年ぶりの鰻温泉に加え、山乃城温泉に今回初めて訪れました。中心には川が流れており、その周囲約100m四方に渡って80°Cを超える熱水がいたるところから湧き出る、壮大な環境でした。天候にも恵まれて予定通りサンプルを採取でき、成果が期待されます。


- 2016.11.02
-
2016年10月21~22日、神奈川県横須賀市JAMSTEC横浜本部において、「ファージ・環境ウイルス研究合同シンポジウム」が開催され、当研究室から2つの口頭発表が行なわれました。
口頭発表 題目西村陽介 環境ウイルスゲノムの自動分類法 綿井博康 Diversity of the Bacteroidetes viruses through analysis of 48 new genera 2016年10月22~25日、神奈川県横須賀市文化会館において、「日本微生物生態学会 第31回大会」が開催され、当研究室から4つのポスター発表が行なわれました。また、当学会のシンポジウムにおいて吉田先生が講演を行いました。
ポスター発表 題目西村陽介 あなたのウイルスゲノムを分類します 綿井博康 genomic-OTUを用いたメタゲノム解析による海洋ウイルスの季節変動 弓矢真穂 超好熱古細菌Aeropyrum pernixに感染する新規溶原化ウイルスの増殖様式に関する一考 大黒達希 水素生成型一酸化炭素資化性好熱菌の分離およびゲノム解析 講演題目
吉田天士 ウイルスと微生物の競合的共進化 また懇親会では、当研究室のOBや関係者と歓談し、産業技術総合研究所の米田恭子博士、京都大学の緒方博之先生と記念写真を撮影しました。とても賑やかで楽しい懇親会となりました。

- 2016.11.01
-
10月16日に、チリのアントファガスタ大学教授のカルロス サラマンカ先生が、3人の研究者らと来研されました。チリでの微細藻利用と魚介類の養殖に関する最近の研究開発事業について紹介があり、澤山教授も交えて情報交換をしました。彼が学生時代の懐かしい話に大変盛り上がりました。カルロス先生はチリで大変活躍をされている有名な先生で、益々のご活躍をお祈りいたします。

- 2016.10.06
-
2016年9月29日、ノルウェーのベルゲン大学からウイルス学者のRuth-Anne Sandaa教授が来研され、ご自身の研究に関するレクチャーをしていただきました。レクチャー後には、一緒に昼食をとりながら歓談し、Sandaa教授と当研究室の学生、院生、スタッフで記念写真を撮影しました。この後、Sandaa教授と吉田先生、院生数名で京都観光に出かけました。

2016年9月28日、基盤研究(S)の共同研究者である高知大学海洋コア総合研究センターの安田尚登先生のもとへ、今後の研究計画の話し合いを兼ねてご挨拶に伺いました。 安田先生から地質学的側面から海洋コア研究を紹介していただき、今後の研究方針を決めていく上での非常に有益な知識を得ることができました。また、施設見学や試験的なサンプリング等もさせていただき、非常に有意義な時間を過ごすことができました。

- 2016.10.05
-
2016年9月11-16日、京都大学において「Extremophiles2016」が開催されました。当研究室から院生1名がポスター発表を行いました。
ポスター発表 題目福山宥斗 Genome Sequence and Transcriptome Analysis of Hydrogenogenic Carboxydotrophic Bacterium, Carboxydothermus pertinax また、Extremophiles2016に、チリのアントファガスタ大学研究員で本研究室OGのGladys Hayasidaさんが参加しポスター発表を行いました。研究室を訪問して下さったので、彼女が本学の院生時代に一緒だった吉田先生とも話が盛り上がりスタッフと記念写真を撮りました。
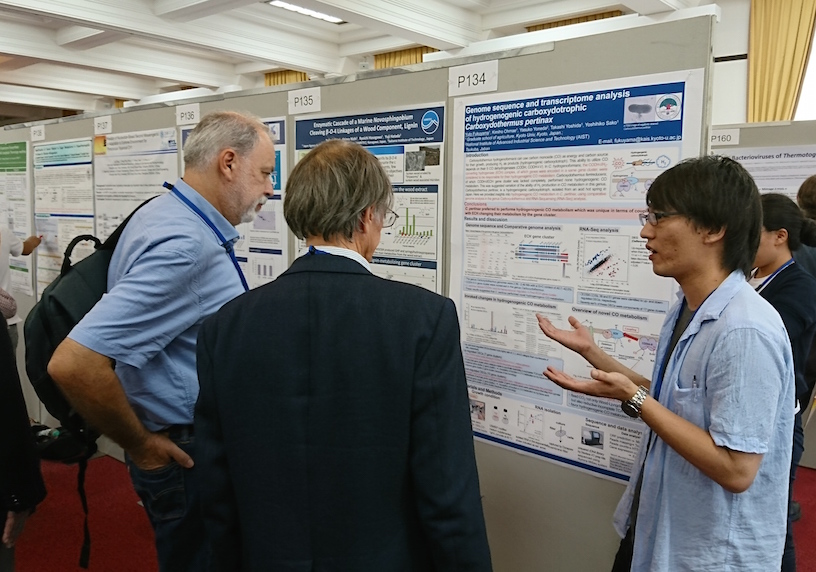

- 2016.08.22
-
2016年8月9,10日、東京大学大気海洋研究所において、「水圏微生物研究フォーラム 2016」が開催されました。当研究室の吉田先生、化学研究所の緒方先生が講演されました。また、当研究室から研究員の西村陽介さん他、院生3名がポスター発表を行いました。
講演題目吉田天士 ウイルスから海洋微生物構造を知る ポスター発表 題目
西村陽介 完全ゲノムから定量化した海洋ウイルスの多様性とその宿主予測 綿井博康 Seasonal dynamics of marine viral communities in Japanese coastal area revealed by genomic OTUs analysis 晴気七菜 大阪湾における微生物群集構造の季節変化 山中ももこ CRISPR配列に基づく有毒ラン藻Microcystis aeruginosaの群集構造解析
- 2016.08.01
-
2016年7月10~13日、英国、プリマスにおいて、「第8回Aquatic virus workshop (8th AVW)」が開催されました。当研究室の吉田先生、西村陽介さん、化学研究所の緒方先生が講演を行いました。
講演題目西村陽介 Metagenomic Whole Genomes of Marine DNA Viruses 吉田天士 Diurnal Dynamics of Novel Viruses And Their Transcription in a Japanese coastal sea by Read Mapping Analyses 本Workshopは参加人数は80名ほどの小規模ですが、著名な研究者が一堂に集まりました。参加者は朝食から夕食まで顔を突き合わせてウイルスのことを語り合うことができました。「ウイルスの世界へようこそ」の雰囲気があり、次々と優秀な若手がこの分野に集まる機会ともなっています。京都大学から世界に向け、ウイルス生態学の次世代展開へ向け一つの方策を提示できました。

- 2016.06.24
-
今年度新たに日本学術振興会に申請していた、科学研究費基盤研究(A)と(S)の採択が決まりました。まず、基盤研究 (A) が採択されました。また、(A)を包括し発展させた基盤研究 (S)(平成28年~32年度)についても ヒアリングに残り、プレゼンの結果採択が決まりました。研究課題は、「時空間的探索による一酸化炭素資化菌の包括的研究とその応用基盤の構築」です。本結果を受けて、(S)の研究課題のみを遂行することになりました。 そこで本研究のスタートを祝してラボでパーティを行いました。 おめでとうございます。 これまでの成果を活かし、さらなる成果を上げるため一丸となって研究を進めていきましょう!

- 2016.06.07
-
2016年5月28~29日、北海道大学大学院水産科学研究院および函館市国際水産・海洋総合センターにおいて、「第18回マリンバイオテクノロジー学会大会」が開催されました。当研究室から院生1名がポスター発表を行いました。
ポスター発表 題目大前公保 水素生成型新規一酸化炭素資化性好熱細菌Calderihabitans maritimusのゲノム解析
- 2016.04.27
-
2016年4月22日、新たなメンバーの歓迎会を行いました。皆で大変盛り上がり楽 しい時間を共有することができました。

- 2016.04.11
-
新年度の顔合わせを行いました。今年は4回生4人、修士1人、PD研究員1人を新たに迎えました。これで4回生4人、修士課程8人、博士課程4人、PD1人と今年度も賑やかな研究室になりました。これからよろしくお願いします。

- 2016.03.26~30
-
2016年3月26~30日、東京海洋大学(品川キャンパス)において、「平成28年度日本水産学会春季大会」が開催されました。当研究室から院生4名が口頭発表を行いました。
口頭発表 題目福山宥斗 オーミックス解析を用いた一酸化炭素 (CO) 資化性菌の CO による転写変動解析 森本大地 アオコ原因ラン藻Microcystis aeruginosaにおけるファージ感染による転写動態変化の網羅的解析 綿井博康 完全長ゲノムに基づく海洋ウイルスの定量的動態解析とその宿主微生物の推定 弓矢真穂 超好熱古細菌Aeropyrum pernixに感染する新規ウイルスの探索 また、当研究室のOBやOGと歓談し、株式会社生物技研の中野江一郎さん、産業技術総合研究所の米田恭子博士と記念写真を撮影しました。

- 2016.02.22
-
2016年2月17日、3月から就職される博士修了2人、修士修了5人(うち2人は博士課程進学)の7人の送別会と謝恩会を行いました。皆様の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2016.02.12
-
2016年2月10日、課題研究発表会が行われました。当研究室から4名がポスター発表を行いました。
ポスター発表 題目笈西優 陸性熱水環境からの新規超好熱古細菌の分離と生理性状解析 大黒達希 カルデラ湖堆積物からの新規好熱性CO資化菌の分離とゲノム解析 晴気七菜 大阪湾に優占する未知ウイルス-微生物感染系の確立の試み 山中ももこ 有毒ラン藻Microcystis aeruginosaの群集構造解析と本種ファージの探索 2016年2月8日、修士論文発表会が行われました。当研究室から5名が口頭発表を行いました。
口頭発表 題目池田栄太朗 陸上熱水環境由来の新規水素生成型一酸化炭素資化性菌の分離とそのゲノム解析 大前公保 新規一酸化炭素資化性好熱菌のゲノム解析を通じたCO代謝の多様性 藤原慎 ウイルス相互作用によるゲノム多様化の解明に向けた超好熱古細菌Aeropyrum pernixの個体群解析 見世健太 一酸化炭素資化性菌の生態学的機能解明を目的とした微生物群集構造解析 綿井博康 完全長ゲノムに基づく海洋微生物ウイルスの定量的動態解析とその宿主予測の試み
- 2016.02.04
-
2016年2月3日、当ラボで博士研究員をされ、現在はマレーシアサインズ大学のCentre for Chemical Biologyにおいて研究をされている古澤剛博士によるセミナーが行われました。興味深い講演をしていただいたあと、古澤博士と当研究室の学生、院生、スタッフで記念写真を撮影しました。古澤博士、ありがとうございました

- 2016.02.02
-
2016年1月4日、当研究室3代目名誉教授の石田祐三郎先生が、新年のご挨拶に農学研究科を訪問され、ご来研されました。歓談の後、石田先生と当研究室の学生、院生、スタッフで記念写真を撮影しました。石田先生は83歳になられた今もご覧のように大変お元気です。

- 2015.12.25
-
2015年12月21日、忘年会を行いました。今年も一年お疲れさまでした。来年からも頑張りましょう!

- 2015.12.17
-
2015年12月17日、博士課程3回生の橋本怜弥さんの博士論文申請講演会が行われました。
申請講演会 題目橋本怜弥 沿岸海域における窒素固定ラン藻の生態・ゲノム学的研究(Ecological and genomic studies on diazotrophic cyanobacteria in coastal seas)
- 2015.11.22~24
-
2015年11月22~24日、福岡国際会議場において、「第63回ウィルス学会学術集会」が開催されました。市民公開講座において吉田天士先生が講演されました。
講演 題目吉田天士 海はウィルスで満ちている
- 2015.11.16
-
2015年11月16日、かずさアカデミアホールにおいて、「藍藻の分子生物学2015」が開催されました。招待講演において吉田天士先生が講演されました。
講演 題目吉田天士 環境におけるMicrocystis aeruginosaとファージのせめぎ合い また、当研究室から院生2名がポスター発表を行いました。
ポスター発表 題目中村銀士 有毒ラン藻Microcystis aeruginosa感染性ファージの地理的差異 橋本怜弥 海洋性窒素固定ラン藻UCYN-Cの新規分離株のゲノム解読
- 2015.10.29~31
-
2015年10月29~31日、京都大学おうばくプラザにおいて、「生命医薬情報学連合大会 2015 年大会(日本バイオインフォマティクス学会2015年年会)」が開催されました。「細菌群集構造解析のためのバイオインフォマティクス実践演習」のセッションにおいて吉田天士先生が講演されました。
講演 題目吉田天士 完全長ゲノムの再構築による海洋ウイルス生態学の転換
- 2015.10.23
-
2015年10月17~20日、茨城県土浦亀城プラザにおいて、「第7回日本・台湾・韓国国際微生物生態シンポジウムおよび日本微生物生態学会第30回大会合同学会」が開催されました。当研究室から院生4名がポスター発表を行いました。
ポスター発表 題目福山宥斗 RNA-Seq provides insights into the novel carbon monoxide metabolism in Carboxydothermus pertinax 森本大地 Transcriptome analysis of Microcystis aeruginosa during Ma-LMM01 infection 大前公保 Genomic adaptation of the thermophilic hydrogenogenic carboxydotrophic bacterium from marine sediment, Calderihabitans maritimus KKC1, driven by carbon monoxide dehydrogenases 綿井博康 Metagenomic analysis of viral communities in Osaka bay, Japan 綿井博康さんはポスター賞を受賞されました。おめでとうございます。
また懇親会では、当研究室のOBや関係者と歓談し、ルイジアナ州立大学のGary M. King博士、海洋研究開発機構の高井研博士、布浦拓郎博士、ファスマック株式会社の中野江一郎さん、東京工業大学の望月智弘博士、水谷紗弥佳博士、産業技術総合研究所の米田恭子博士、京都大学の中川聡先生、神川龍馬先生、緒方博之先生、西村陽介さんと記念写真を撮影しました。とても賑やかな懇親会となりました。



- 2015.07.10
-
2015年7月10日に当ラボOBで現在和歌山県水産試験場で研究活動を行っている中山仁志博士が来研されました。博士の研究対象であるアユの冷水病についての講演をしていただき、和歌山県の水産環境やアユとその感染症について大変勉強になりました。先輩から大いに刺激を受け、有意義な時間となりました。中山博士、ありがとうございました。
- 2015.06.26
-
2015年6月26日に、京都大学化学研究所バイオインフォマティクスセンター緒方研究室の客員准教授のPascal Hingamp博士によるランチョンセミナーが行われました。皆でピザランチやお話を楽しんだ後、Pascal博士が参加するタラ海洋探査国際・学際コンソーシアムによる研究成果について、興味深い講演をしていただきました。この研究成果は、既に米国科学誌Scienceに5報の論文として掲載されており、海洋性浮遊生物に関する地球規模での研究知見に、大変刺激を受けました。Pascal博士、ありがとうございました。
講演者 題目
Dr. Pascal Hingamp Tara Oceans Project: marine plankton at planetary scale
- 2015.06.07
-
2015年6月7日~11日にオランダで開催されたThe 6th Congress of European Microbiologists (FEMS 2015) で、当ラボの吉田天士准教授が発表しました。
題目Population dynamics of a cyanobacterium microcystis aeruginosa using amplicon analysis for the leader-end crispr fragments
全体として、医学系や食品系の発表が多く、ヨーロッパにおいても環境におけるウイルスの認知度はまだまだ低いようです。しかしながら、大会のオープニングレクチャーは環境ウイルスの数の多さや多様性に注目したものであり、今後環境ウイルス学の益々の発展が期待されるとのことでした。
- 2015.06.02
-
2015年6月2日に、チリのアントファガスタ大学 海洋科学部教授のカルロス リケルメ博士が来研されました。 カルロス先生は本研究室で博士を取得して、母校のアントファガスタ大学で教授となられ、海洋微細藻が生産する抗酸化作用を有するルテインの効率的な生産系に
ついて研究されています。久しぶりに研究や留学生の受け入れ等について、楽しく懐かしい話が弾みました。
お元気で益々のご活躍をお祈りいたします。

- 2015.06.01
-
2015年5月30、31日、東京海洋大学において第17回日本マリンバイオテクノロジー学会大会が開催されました。当研究室から2名が口頭発表を行いました。
口頭発表 題目藤原慎 超好熱古細菌Aeropyrum属における宿主-ウイルス相互作用の解明 弓矢真穂 超好熱古細菌Aeropyrum pernixに感染する新規ウイルスの探索 写真は口頭発表後、東京海洋大学キャンパス内にて撮影しました。

- 2015.05.16
-
新入生歓迎会を行いました。皆で楽しい一時を過ごすことができました。

- 2015.04.21
-
新年度の顔合わせを行いました。今年は四回生四人、博士一人を新たに迎えました。これからよろしくお願いします。

- 2015.04.15
-
2015年3月27日~31日、東京海洋大学において平成27年度日本水産学会春季大会が開催されました。当研究室から6名が口頭発表を行いました。 口頭発表 題目
中村銀士 共存する細菌・ファージ感染系によるラン藻Mycrocystis aeruginosaの間接的な溶藻 橋本怜弥 海洋性新規窒素固定ラン藻YR-1株のゲノム解読 福山宥斗 網羅的転写解析による一酸化炭素(CO)資化性菌のCO代謝機構の解明 大前公保 水素生成型一酸化炭素資化性菌Calderihabitans maritimus KKC1と酢酸生成菌Moorella spp.の比較ゲノム解析 見世健太 次世代シーケンサーを用いた高温強酸温泉における微生物群集構造解析 綿井博康 次世代シーケンサーを用いた未知の海洋細菌-ウイルス感染系の推定 吉田天士先生が平成26年度日本水産学会進歩賞を受賞されました。受賞式のあと、東京海洋大学の桜の下で記念写真を撮りました。吉田先生、おめでとうございます。

また会員交歓会では、当研究室のOBや関係者と歓談し、福井県立大学の横山芳博先生と前田知己さん、近畿大学の安藤正史先生、東京工業大学の望月智弘さんと記念写真を撮りました。とても賑やかな交歓会となりました。

- 2015.03.16
-
2015年2月26日、3月から就職されるPD1人、博士修了1人、修士修了3人(うち1人は博士課程進学)の4人の送別会と謝恩会を行いました。先輩方の益々のご活躍とご多幸を心よりお祈り申し上げます。

- 2015.02.13
-
2015年2月13日、卒業論文のポスター発表が行われました。当研究室から4名が発表しました。口頭発表 題目
清水千尋 熱水環境由来の水素生成型CO資化集積培養系のゲノム解析 中田舞 Microcystis aeruginosaにおけるファージ耐性に関与する遺伝子変異の探索 弓矢真穂 超好熱古細菌Aeropyrum pernixに感染する新規ウィルスの分離と性状解析 吉安勇人 未知の宿主-ウィルス感染系探索のためのMagneto-FISH法の確立
- 2015.02.12
-
2015年2月9日、修士論文の口頭発表が行われました。当研究室から3名が発表しました。口頭発表 題目
上原美翔 有毒ラン藻Microcystis aeruginosaにおけるファージ耐性遺伝子領域の多様化 福山宥斗 好熱細菌Carboxydothermus pertinax Ug1におけるCOの酸化と共役した新規水素生成経路の解明 増田真帆 新規系統群に属する海産窒素固定ラン藻のゲノム解析
- 2015.02.09
-
2015年1月20~22日、伊豆半島の熱水環境においてサンプリングを行いました。多様な泉源をめぐることができ、充実したサンプリングとなりました。


- 2014.12.26
-
2014年12月26日、大掃除と忘年会を行いました。2015年が素敵な年となりますように。また、吉田天士先生が3月に日本水産学会進歩賞を受賞されることが決まり、ケーキでお祝いをしました。吉田先生、おめでとうございます。


- 2014.12.05
-
2014年11月22日、京都大学において日本水産学会近畿支部例会が開催されました。当研究室から2名が口頭発表を行いました。
口頭発表 題目大福高史 好熱古細菌クレンアーキオータ門のゲノム進化 福山宥斗 好熱菌Carboxydothermus pertinaxの一酸化炭素代謝と共役する新規水素生成機構の探索 当口頭発表において福山さんは優秀発表賞を受賞されました。おめでとうございます。
- 2014.11.20
-
2014年11月20日、博士課程3回生の大福高史さんの博士論文申請講演会が行われました。
申請講演会 題目大福高史 Genomic analysis of the marine hyperthermophilic archaeon Aeropyrum (海洋性超好熱古細菌Aeropyrum属のゲノム解析)
- 2014.11.17
-
2014年10月21~24日、アクトシティ浜松において環境微生物系学会合同大会2014が開催されました。当研究室から2名がポスター発表、2名が口頭発表を行いました。 口頭発表 題目
上原美翔 環境中におけるラン藻 Microcystis aeruginosa の CRISPR に基づく個体群解析 藤原慎 超好熱古細菌 Aeropyrum 属におけるウィルス感染履歴配列の解析 ポスター発表 題目
本田貴史 ファージ Ma-LMM01 感染過程における Microcystis aeruginosa の遺伝子転写解析 池田栄太朗 熱水環境中の一酸化炭素資化性好熱菌の分子生態学的研究 写真はポスター発表会場にて当研究室のOB、OGと撮影しました。

- 2014.10.16
-
2014年9月20~21日、九州大学において平成26年度日本水産学会秋季大会が開催されました。当研究室から3名が口頭発表を行いました。 口頭発表 題目
橋本怜弥 瀬戸内海における単細胞性窒素固定ラン藻の生態学的研究 綿井博康 日本近海における共生型単細胞性窒素固定ラン藻とその宿主藻類の生態学的研究 大前公保 海洋性水素生成型一酸化炭素資化性好熱細菌Calderihabitans maritimus KKC1のゲノム解析 また会員交歓会では、当研究室のOBや関係者と歓談し記念写真を撮りました。

- 2014.06.27
-
キャノン財団理想の追求「ウイルスは海洋生物多様性を創生・維持する素粒子か?」のキックオフミーティングを、京大化学研究所バイオインフォマティクスセンター(緒方教授、五斗准教授)にて、多くの両院生が参加して開催しました。写真は、お世話になっているスーパーコンピューターです。

- 2014.05.27-29
-
修士課程の4人が九州の温泉環境でサンプリングを行ってきました。充実したサンプリングでした。


- 2014.04.23
-
新入生歓迎会を行いました。みんなで楽しい時間を過ごすことができました。

- 2014.04.18
-
東京都大手町にある経団連にて、キヤノン財団贈呈式が行われました。研究代表者の吉田先生が、キヤノン(株) CEOの御手洗富士夫会長から研究費を晴れて授与されました。

- 2014.04.04
-
新年度の顔合わせを行いました。今年は新たに4回生4人、修士1回生1人を迎えました。これからよろしくお願いします。

- 2014.02.19
-
修士論文の口頭発表が行われました。当研究室から2名が発表しました。
高尾郷介 一酸化炭素資化性好熱菌Carboxydothermus属の比較ゲノム解析 高橋春菜 有毒ラン藻Microcystis aeruginosaファージの感染過程における転写制御機構に関する研究 また、同日、送別会を行いました。先輩方の今後のご活躍を心よりお祈り申し上げます。

- 2014.02.12
-
卒業論文のポスター発表が行われました。当研究室から4名が発表しました。
池田栄太朗 熱水環境における一酸化炭素資化性菌の生態学的研究 大前公保 CO資化性好熱菌Calderihabitans maritimus KKC1株のゲノム解析 藤原慎 超好熱古細菌Aeropyrum属におけるウイルス感染履歴配列の解析 綿井博康 日本近海における共生型単細胞性窒素固定ラン藻の分離およびその宿主藻類の生態学的研究
- 2014.02.12
-
卒業論文のポスター発表が行われました。当研究室から4名が発表しました。
池田栄太朗 熱水環境における一酸化炭素資化性菌の生態学的研究 大前公保 CO資化性好熱菌Calderihabitans maritimus KKC1株のゲノム解析 藤原慎 超好熱古細菌Aeropyrum属におけるウイルス感染履歴配列の解析 綿井博康 日本近海における共生型単細胞性窒素固定ラン藻の分離およびその宿主藻類の生態学的研究
- 2014.01.23
-
2014年1月16日、博士課程3回生の本田貴史さん、佐藤希美さんが博士論文申請講演会を行われました。
本田貴史 Studies on transcriptional regulation in a toxic cyanobacterium Microcystis aeruginosa
(有毒ラン藻 Microcystis aeruginosa の遺伝子転写制御に関する研究)佐藤希美 X-ray crystal structure analysis of reduced and ligand-bound nitric oxide reductase from Pseudomonas aeruginosa
(Pseudomonas aeruginosa 由来還元型および配位子結合型一酸化窒素還元酵素のX線結晶構造解析)
- 2013.12.27
-
2013年12月26日、大掃除と忘年会を行いました。
今年も一年お疲れさまでした。来年からも頑張りましょう!
- 2013.11.29
-
2013年11月23~25日、鹿児島大学において第29回日本微生物生態学会大会が開催されました。 当ラボからも3つの口頭発表、2つのポスター発表を行いました。
口頭発表 題目橋本怜弥 瀬戸内海における単細胞性窒素固定ラン藻の分布調査 福山宥斗 一酸化炭素資化性好熱菌Carboxydothermus pertinax のCO代謝 米田恭子 熱水環境中の一酸化炭素資化性好熱Carboxydothermusの分子生態学的研究 ポスター発表 題目
中村銀士 有毒ラン藻Microcystis aeruginosa感染性シアノファージの遺伝的多様性 大福高史 超好熱アーキアAeropyrum属のゲノム多様性
当ポスター発表において、中村さんは優秀ポスター賞を受賞されました。おめでとうございます。
- 2013.11.21
-
2013年11月21日、博士課程3回生の米田恭子さん、井上喬裕さんが博士論文申請講演会を行われました。
米田恭子 Physiological and ecological studies on novel _carboxydotrophic thermophiles (新規一酸化炭素資化性好熱菌の生理生態学的研究) 井上喬裕 Genetic engineering studies of Ni-carbon monoxide _dehydrogenase from a thermophilic carboxydotrophic _bacterium (好熱性カルボキシドトロフ由来一酸化炭素デヒドロゲナーゼに関する遺伝子工学的研究)
- 2013.11.03~07
-
2013年11月3~7日、アメリカのフロリダでAquatic Virus Workshop 7が開催されました。当ラボからは次の口頭発表を行いました。
吉田 天士 Co-evolution between the bloom-forming cyanobacteria Microcystis aeruginosa and its phages 
当研究室に興味を持たれた方へ
当研究室に興味を持たれた方はこちらをご覧ください
所在地・連絡先
〒606-8502 京都市左京区北白川追分町
京都大学大学院農学研究科
応用生物科学専攻海洋分子微生物学分野
農学部総合館 4階 E410号室
Tel; 075-753-6217
Fax; 075-753-6226
e-mail; yoshida.takashi.7a * kyoto-u.ac.jp (* を@としてください)
Firefox上では本ページの画面が正しく表示されないことがあるようです。その際は他のブラウザで閲覧ください。